

Downregulation of pathways implicated in liver inflammation and tumorigenesis of glycogen storage disease type Ia mice receiving gene therapy
- PMID: 28334808
- PMCID: PMC6075378
- DOI: 10.1093/hmg/ddx097
Downregulation of pathways implicated in liver inflammation and tumorigenesis of glycogen storage disease type Ia mice receiving gene therapy
Abstract
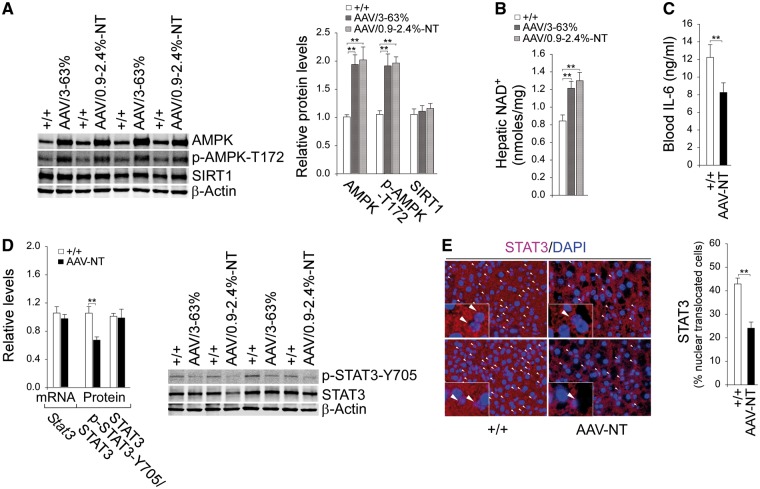
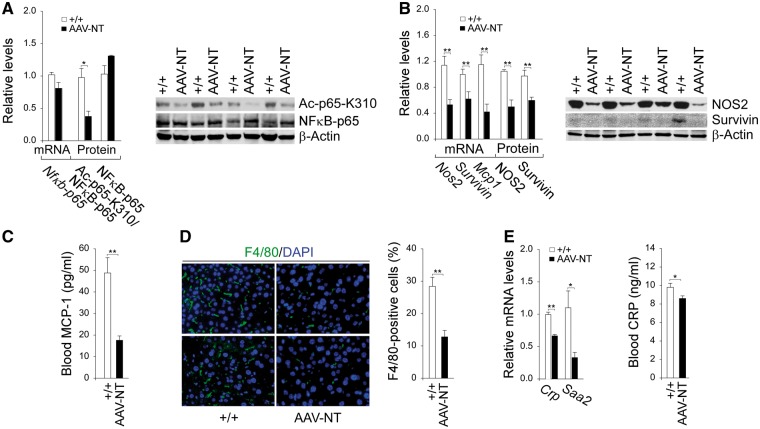
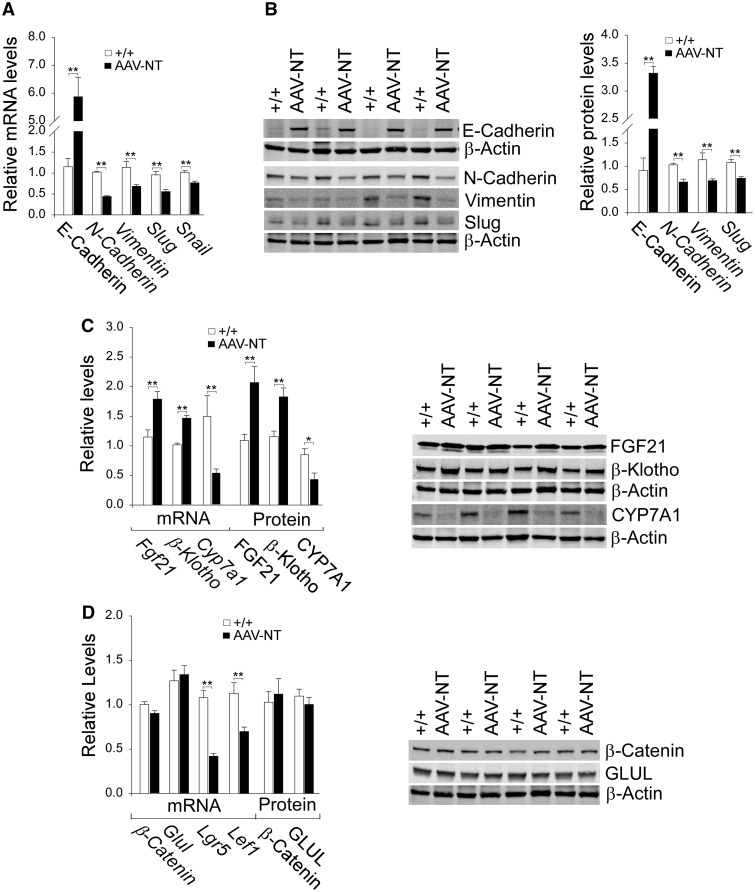
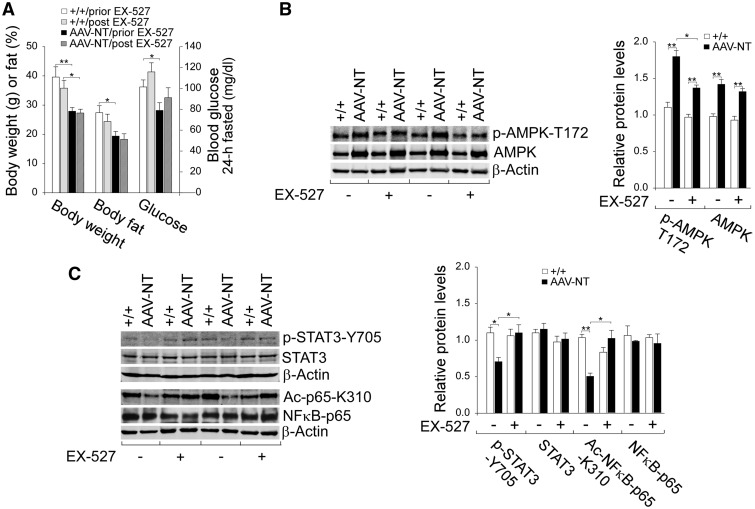
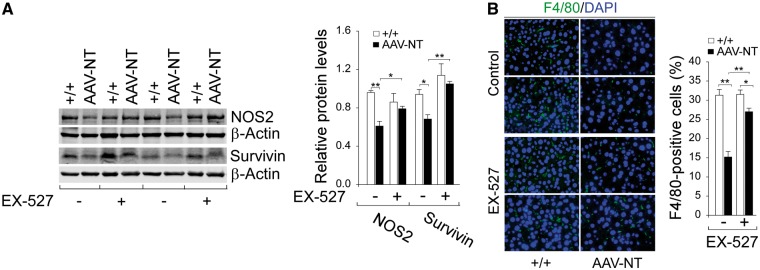
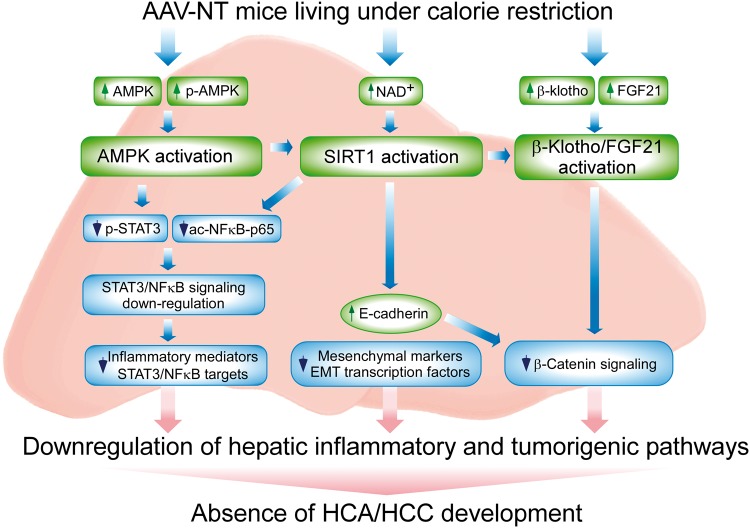
Glycogen storage disease type Ia (GSD-Ia) is characterized by impaired glucose homeostasis and long-term risks of hepatocellular adenoma (HCA) and carcinoma (HCC). We have shown that the non-tumor-bearing (NT), recombinant adeno-associated virus (rAAV) vector-treated GSD-Ia mice (AAV-NT mice) expressing a wide range (0.9-63%) of normal hepatic glucose-6-phosphatase-α activity maintain glucose homeostasis and display physiologic features mimicking animals living under calorie restriction (CR). We now show that in AAV-NT mice, the signaling pathways of the CR mediators, AMP-activated protein kinase (AMPK) and sirtuin-1 are activated. AMPK/sirtuin-1 inhibit the activity of STAT3 (signal transducer and activator of transcription 3) and NFκB (nuclear factor κB), the pro-inflammatory and cancer-promoting transcription factors. Sirtuin-1 also inhibits cancer metastasis via increasing the expression of E-cadherin, a tumor suppressor, and decreasing the expression of mesenchymal markers. Consistently, in AAV-NT mice, hepatic levels of active STAT3 and NFκB-p65 were reduced as were expression of mesenchymal markers, STAT3 targets, NFκB targets and β-catenin targets, all of which were consistent with the promotion of tumorigenesis. AAV-NT mice also expressed increased levels of E-cadherin and fibroblast growth factor 21 (FGF21), targets of sirtuin-1, and β-klotho, which can acts as a tumor suppressor. Importantly, treating AAV-NT mice with a sirtuin-1 inhibitor markedly reversed many of the observed anti-inflammatory/anti-tumorigenic signaling pathways. In summary, activation of hepatic AMPK/sirtuin-1 and FGF21/β-klotho signaling pathways combined with down-regulation of STAT3/NFκB-mediated inflammatory and tumorigenic signaling pathways can explain the absence of hepatic tumors in AAV-NT mice.
Published by Oxford University Press 2017. This work is written by US Government employees and is in the public domain in the US.
Figures

References
-
- Rake J.P., Visser G., Labrune P., Leonard J.V., Ullrich K., Smit G.P. (2002) Glycogen storage disease type I: diagnosis, management, clinical course and outcome. Results of the European Study on Glycogen Storage Disease Type I (ESGSD I). Eur. J. Pediatr., 161 (Suppl 1), S20–S34. - PubMed
-
- Greene H.L., Slonim A.E., O’Neill J.A. Jr, Burr I.M. (1976) Continuous nocturnal intragastric feeding for management of type 1 glycogen-storage disease. N. Engl. J. Med., 294, 423–425. - PubMed
-
- Chen Y.T., Cornblath M., Sidbury J.B. (1984) Cornstarch therapy in type I glycogen-storage disease. N. Engl. J. Med., 310, 171–175. - PubMed
-
- Franco L.M., Krishnamurthy V., Bali D., Weinstein D.A., Arn P., Clary B., Boney A., Sullivan J., Frush D.P., Chen Y.T.. et al. (2005) Hepatocellular carcinoma in glycogen storage disease type Ia: a case series. J. Inherit. Metab. Dis., 28, 153–162. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
