

Aicardi-Goutières syndrome protein TREX1 suppresses L1 and maintains genome integrity through exonuclease-independent ORF1p depletion
- PMID: 28334850
- PMCID: PMC5416883
- DOI: 10.1093/nar/gkx178
Aicardi-Goutières syndrome protein TREX1 suppresses L1 and maintains genome integrity through exonuclease-independent ORF1p depletion
Abstract
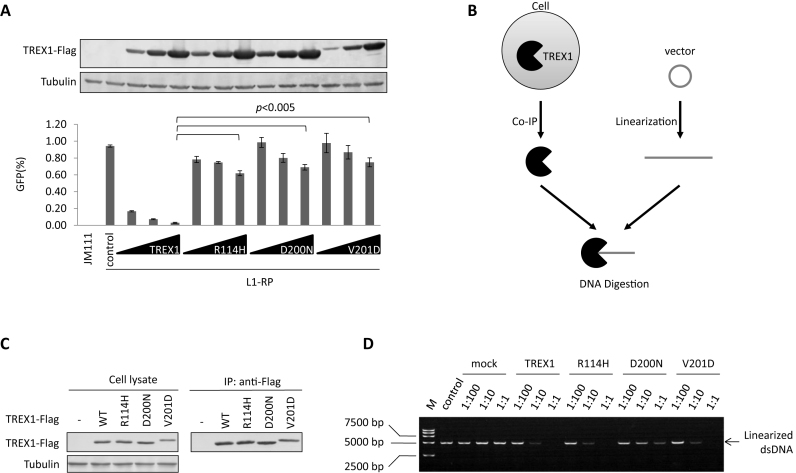
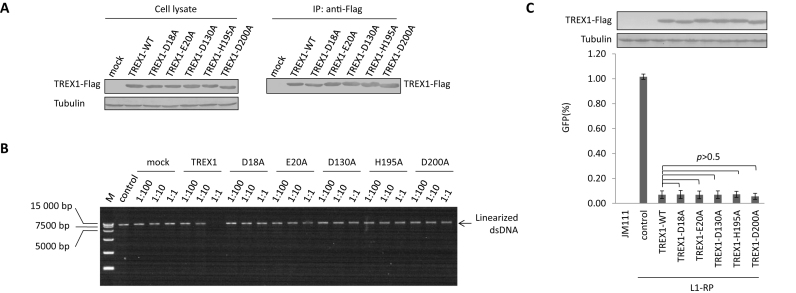
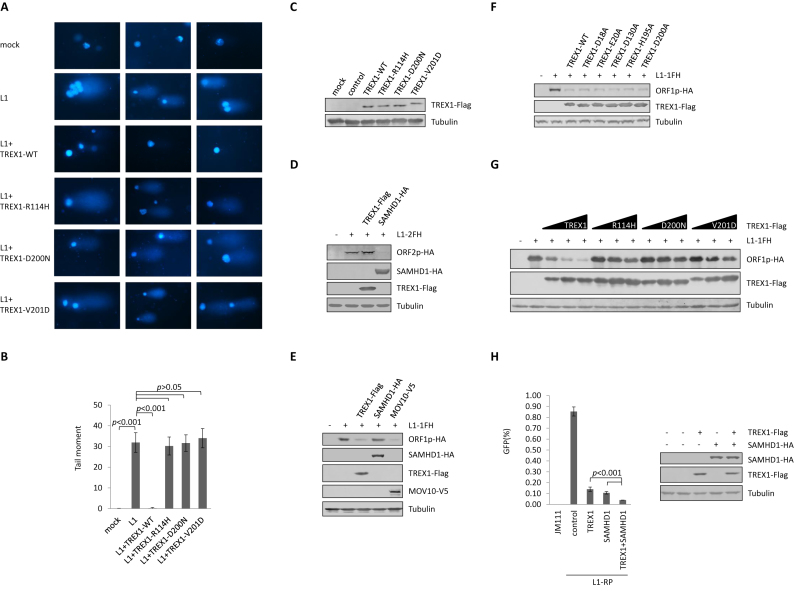
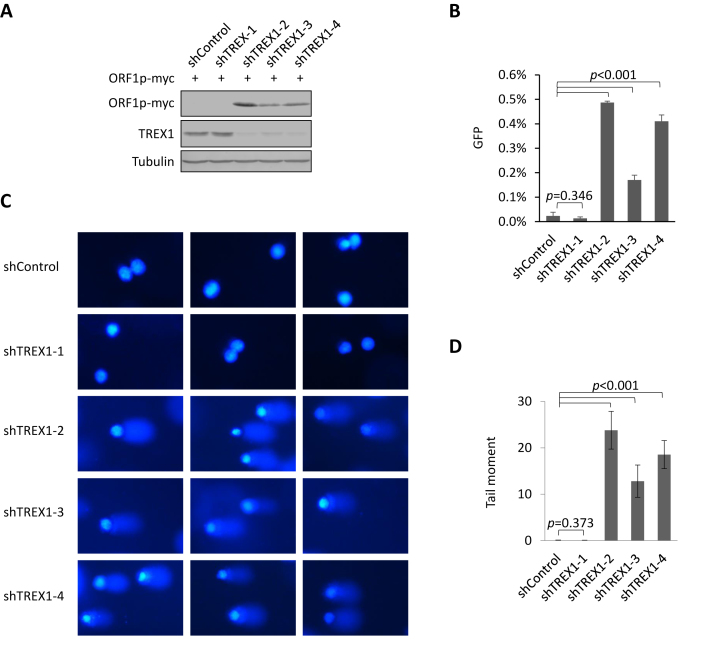
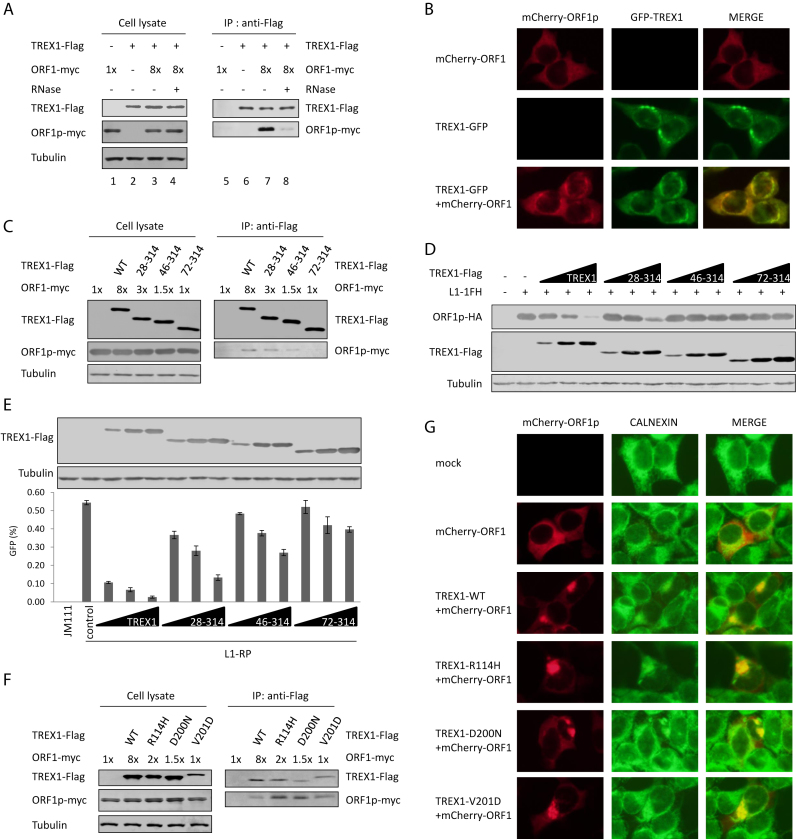
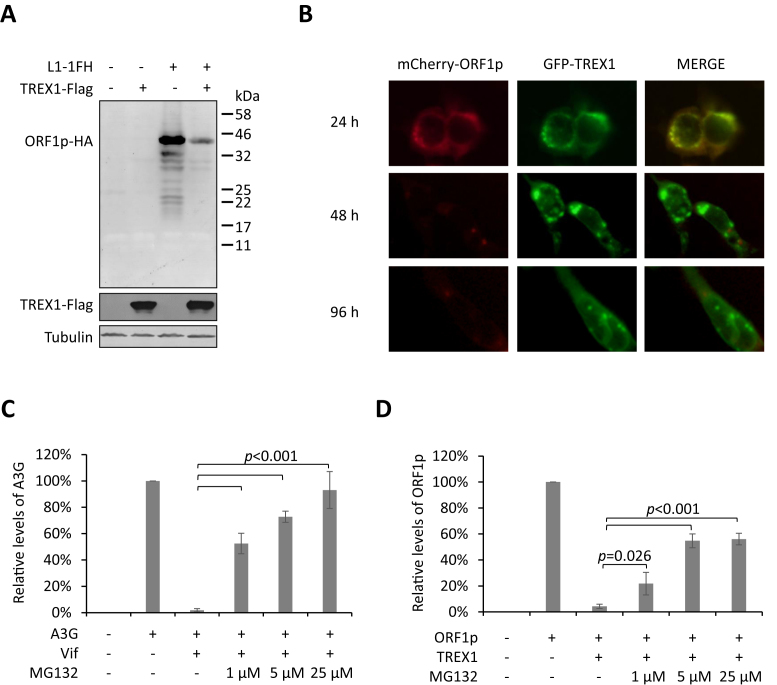
Maintaining genome integrity is important for cells and damaged DNA triggers autoimmunity. Previous studies have reported that Three-prime repair exonuclease 1(TREX1), an endogenous DNA exonuclease, prevents immune activation by depleting damaged DNA, thus preventing the development of certain autoimmune diseases. Consistently, mutations in TREX1 are linked with autoimmune diseases such as systemic lupus erythematosus, Aicardi-Goutières syndrome (AGS) and familial chilblain lupus. However, TREX1 mutants competent for DNA exonuclease activity are also linked to AGS. Here, we report a nuclease-independent involvement of TREX1 in preventing the L1 retrotransposon-induced DNA damage response. TREX1 interacted with ORF1p and altered its intracellular localization. Furthermore, TREX1 triggered ORF1p depletion and reduced the L1-mediated nicking of genomic DNA. TREX1 mutants related to AGS were deficient in inducing ORF1p depletion and could not prevent L1-mediated DNA damage. Therefore, our findings not only reveal a new mechanism for TREX1-mediated L1 suppression and uncover a new function for TREX1 in protein destabilization, but they also suggest a novel mechanism for TREX1-mediated suppression of innate immune activation through maintaining genome integrity.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Gehrke N., Mertens C., Zillinger T., Wenzel J., Bald T., Zahn S., Tuting T., Hartmann G., Barchet W.. Oxidative damage of DNA confers resistance to cytosolic nuclease TREX1 degradation and potentiates STING-dependent immune sensing. Immunity. 2013; 39:482–495. - PubMed
-
- Luan D.D., Korman M.H., Jakubczak J.L., Eickbush T.H.. Reverse transcription of R2Bm RNA is primed by a nick at the chromosomal target site: a mechanism for non-LTR retrotransposition. Cell. 1993; 72:595–605. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
