

A class of circadian long non-coding RNAs mark enhancers modulating long-range circadian gene regulation
- PMID: 28335007
- PMCID: PMC5449593
- DOI: 10.1093/nar/gkx156
A class of circadian long non-coding RNAs mark enhancers modulating long-range circadian gene regulation
Abstract
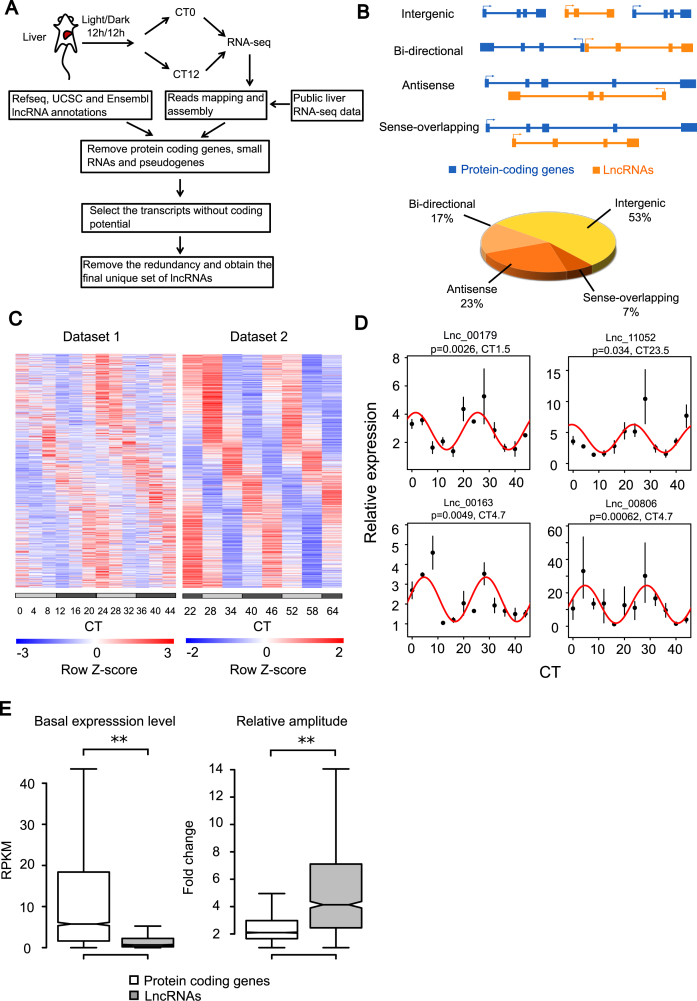
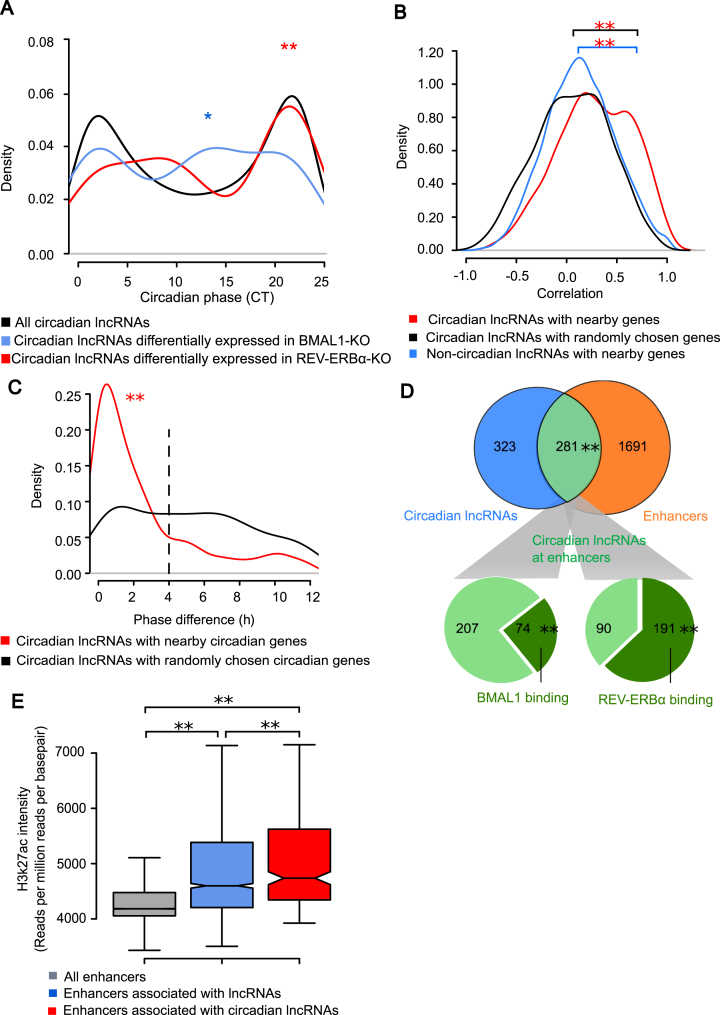
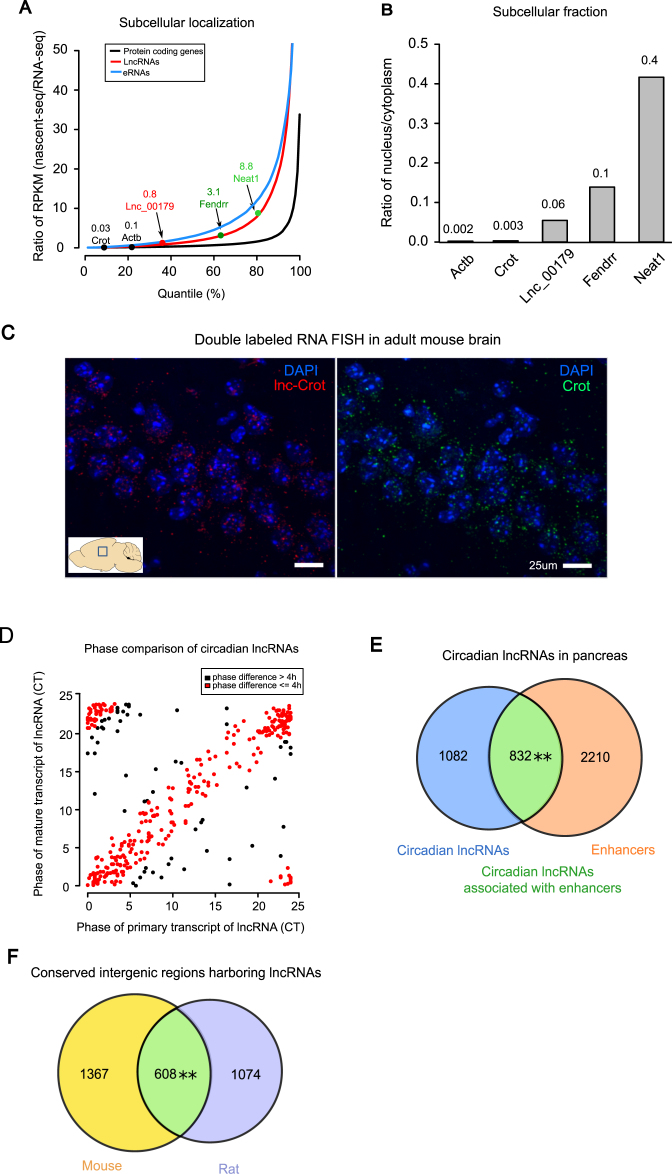
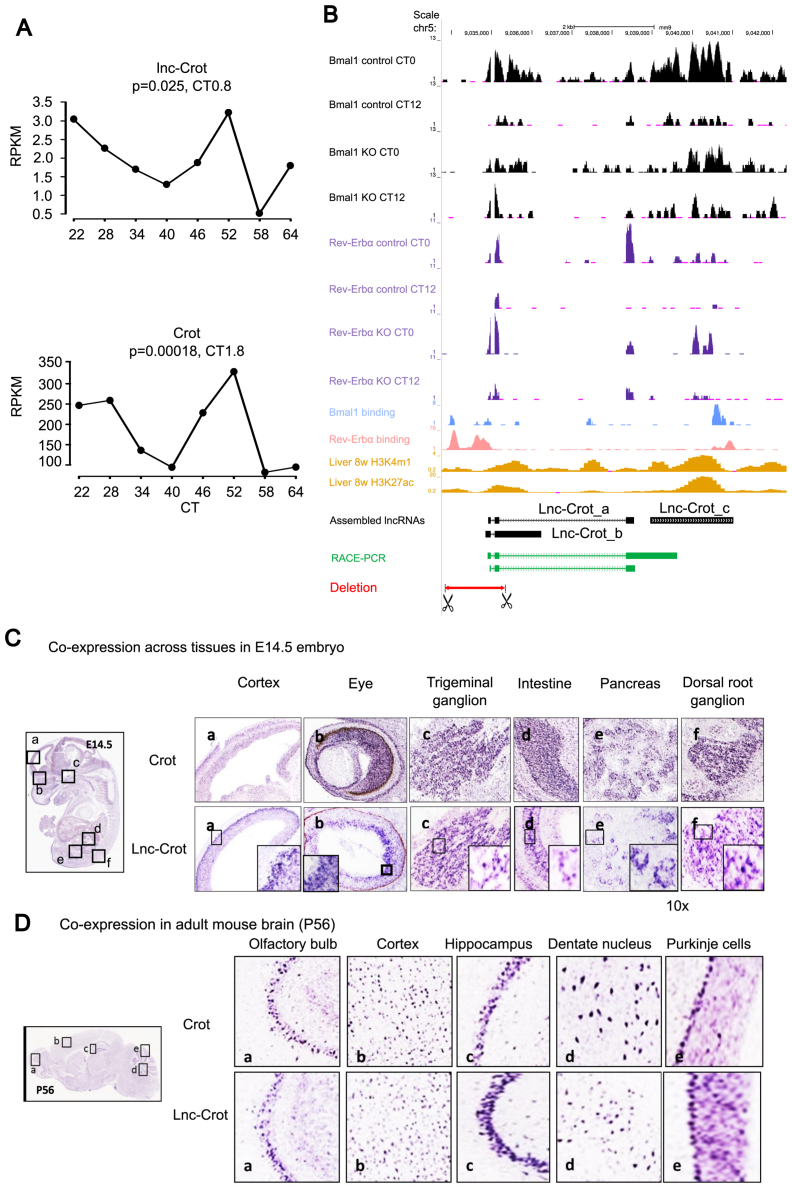
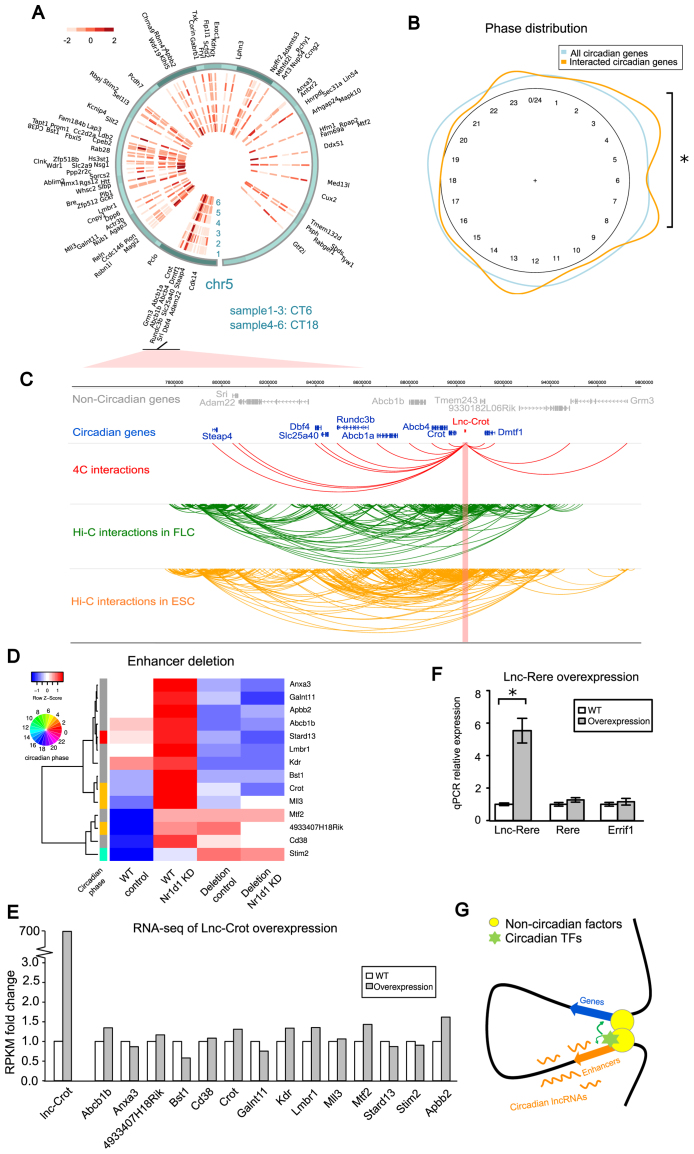
Circadian rhythm exerts its influence on animal physiology and behavior by regulating gene expression at various levels. Here we systematically explored circadian long non-coding RNAs (lncRNAs) in mouse liver and examined their circadian regulation. We found that a significant proportion of circadian lncRNAs are expressed at enhancer regions, mostly bound by two key circadian transcription factors, BMAL1 and REV-ERBα. These circadian lncRNAs showed similar circadian phases with their nearby genes. The extent of their nuclear localization is higher than protein coding genes but less than enhancer RNAs. The association between enhancer and circadian lncRNAs is also observed in tissues other than liver. Comparative analysis between mouse and rat circadian liver transcriptomes showed that circadian transcription at lncRNA loci tends to be conserved despite of low sequence conservation of lncRNAs. One such circadian lncRNA termed lnc-Crot led us to identify a super-enhancer region interacting with a cluster of genes involved in circadian regulation of metabolism through long-range interactions. Further experiments showed that lnc-Crot locus has enhancer function independent of lnc-Crot's transcription. Our results suggest that the enhancer-associated circadian lncRNAs mark the genomic loci modulating long-range circadian gene regulation and shed new lights on the evolutionary origin of lncRNAs.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Thresher R.J., Vitaterna M.H., Miyamoto Y., Kazantsev a, Hsu D.S., Petit C., Selby C.P., Dawut L., Smithies O., Takahashi J.S. et al. Role of mouse cryptochrome blue-light photoreceptor in circadian photoresponses. Science. 1998; 282:1490–1494. - PubMed
-
- Preitner N., Damiola F., Zakany J., Duboule D., Albrecht U., Schibler U.. The orphan nuclear receptor REV-ERBα controls circadian transcription within the positive limb of the mammalian circadian oscillator University of Geneva University of Geneva. Cell. 2002; 110:251–260. - PubMed
-
- Panda S., Antoch M.P., Miller B.H., Su A.I., Schook A.B., Straume M., Schultz P.G., Kay S.A., Takahashi J.S., Hogenesch J.B.. Coordinated transcription of key pathways in the mouse by the circadian clock. Cell. 2002; 109:307–320. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
