

Monitoring Cell-surface N-Glycoproteome Dynamics by Quantitative Proteomics Reveals Mechanistic Insights into Macrophage Differentiation
- PMID: 28336715
- PMCID: PMC5417820
- DOI: 10.1074/mcp.M116.063859
Monitoring Cell-surface N-Glycoproteome Dynamics by Quantitative Proteomics Reveals Mechanistic Insights into Macrophage Differentiation
Abstract
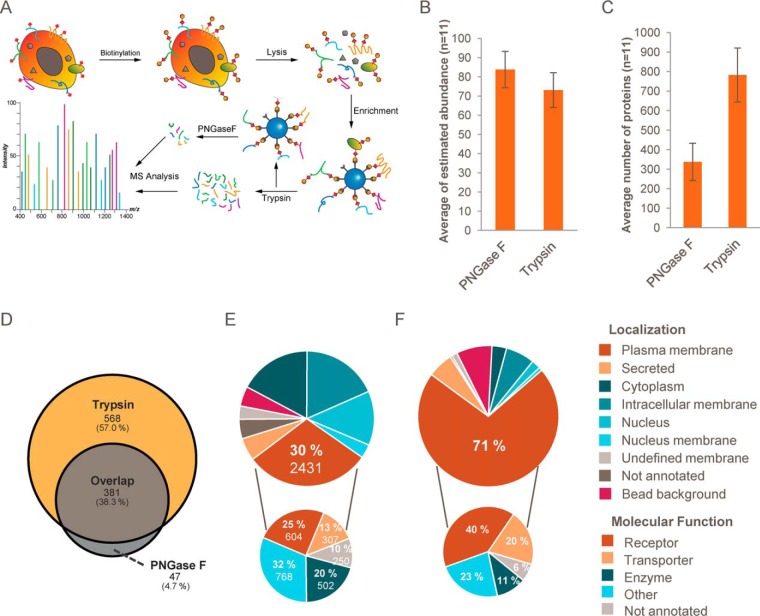
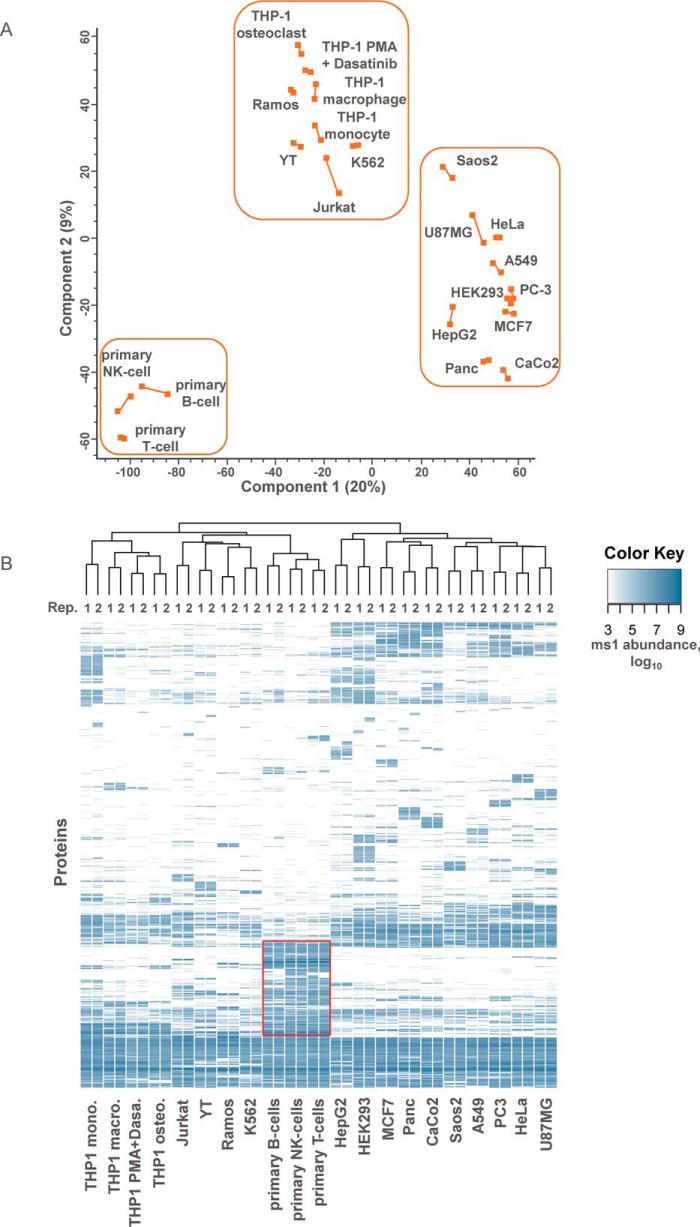
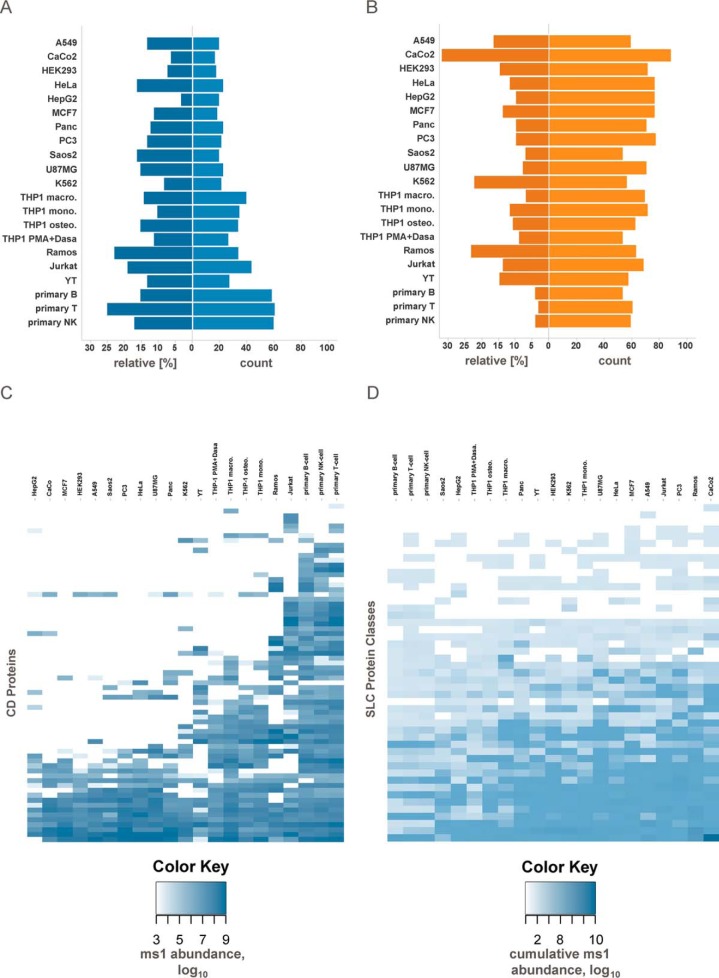
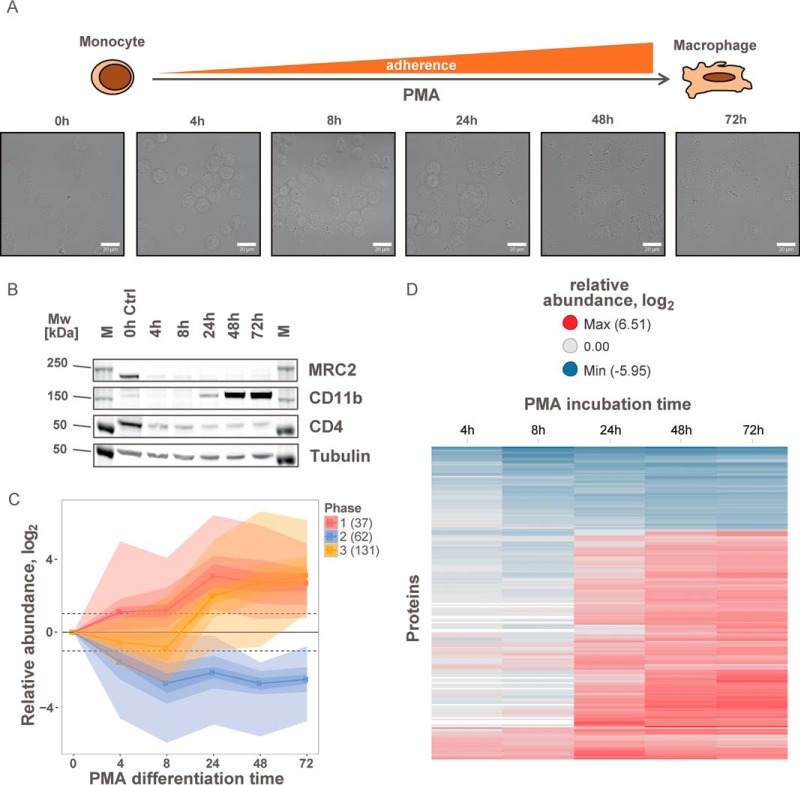
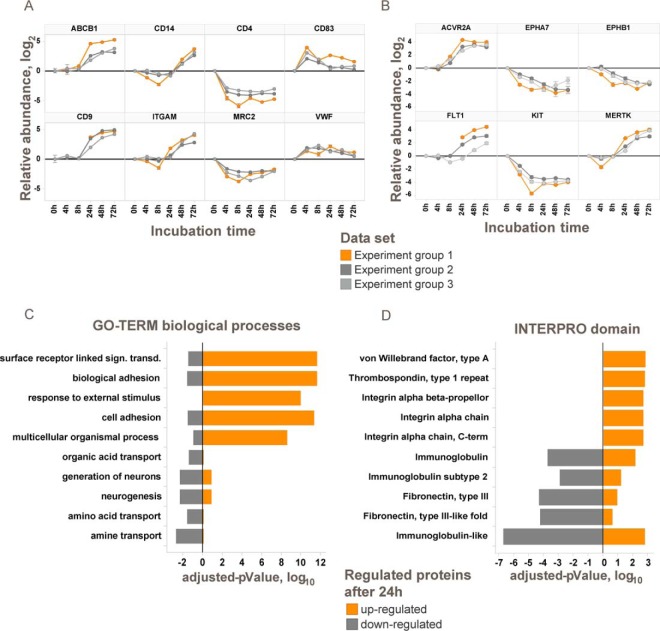
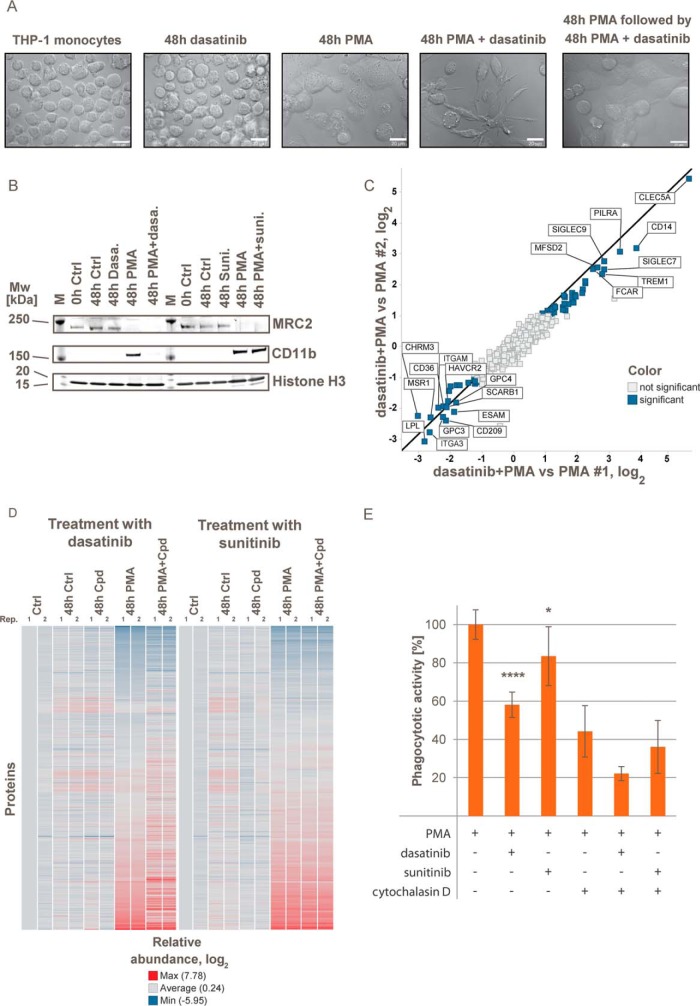
The plasma membrane proteome plays a crucial role in inter- and intracellular signaling, cell survival, and cell identity. As such, it is a prominent target for pharmacological intervention. The relatively low abundance of this subproteome in conjunction with challenging extractability and solubility still hampers its comprehensive analysis. Here, we combined a chemical glycoprotein-tagging strategy with mass spectrometry to enable comprehensive analysis of the cell-surface glycoproteome. To benchmark this workflow and to provide guidance for cell line selection for functional experiments, we generated an inventory of the N-linked cell-surface glycoproteomes of 15 standard laboratory human cell lines and three primary lymphocytic cell types. On average, about 900 plasma membrane and secreted proteins were identified per experiment, including more than 300 transporters and ion channels. Primary cells displayed distinct expression of surface markers and transporters underpinning the importance of carefully validating model cell lines selected for the study of cell surface-mediated processes. To monitor dynamic changes of the cell-surface proteome in a highly multiplexed experiment, we employed an isobaric mass tag-based chemical labeling strategy. This enabled the time-resolved analysis of plasma membrane protein presentation during differentiation of the monocytic suspension cell line THP-1 into macrophage-like adherent cells. Time-dependent changes observed in membrane protein presentation reflect functional remodeling during the phenotypic transition in three distinct phases: rapid surface presentation and secretion of proteins from intracellular pools concurrent with rapid internalization of no longer needed proteins and finally delayed presentation of newly synthesized macrophage markers. Perturbation of this process using marketed receptor tyrosine kinase inhibitors revealed dasatinib to severely compromise macrophage differentiation due to an off-target activity. This finding suggests that dynamic processes can be highly vulnerable to drug treatment and should be monitored more rigorously to identify adverse drug effects.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
All authors are employees of this company
Figures
References
-
- Anderle P., Huang Y., and Sadée W. (2004) Intestinal membrane transport of drugs and nutrients: genomics of membrane transporters using expression microarrays. Eur. J. Pharm. Sci. 21, 17–24 - PubMed
-
- Navarro G., Cordomí A., Zelman-Femiak M., Brugarolas M., Moreno E., Aguinaga D., Perez-Benito L., Cortés A., Casadó V., Mallol J., Canela E. I., Lluís C., Pardo L., García-Sáez A. J., McCormick P. J., and Franco R. (2016) Quaternary structure of a G-protein-coupled receptor heterotetramer in complex with Gi and Gs. BMC Biol. 14, 26. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
