

Review
doi: 10.3389/fimmu.2017.00247.
eCollection 2017.
Nutrient and Metabolic Sensing in T Cell Responses
Affiliations
- PMID: 28337199
- PMCID: PMC5343023
- DOI: 10.3389/fimmu.2017.00247
Item in Clipboard
Review
Nutrient and Metabolic Sensing in T Cell Responses
Front Immunol.
.
Abstract
T cells play pivotal roles in shaping host immune responses in infectious diseases, autoimmunity, and cancer. The activation of T cells requires immune and growth factor-derived signals. However, alterations in nutrients and metabolic signals tune T cell responses by impinging upon T cell fates and immune functions. In this review, we summarize how key nutrients, including glucose, amino acids, and lipids, and their sensors and transporters shape T cell responses. We also briefly discuss regulation of T cell responses by oxygen and energy sensing mechanisms.
Keywords: T cell response; amino acid; energy; glucose; lipid; oxygen.
Figures
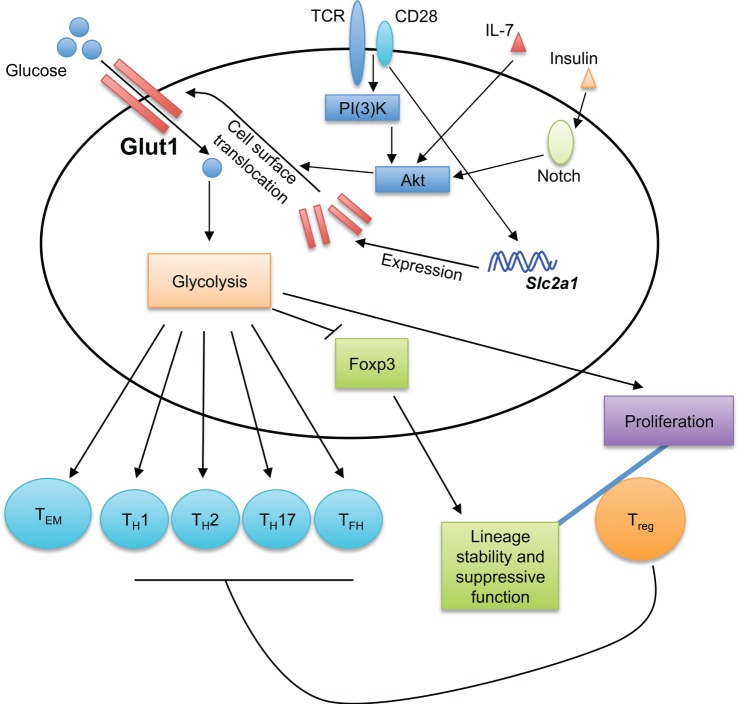
Glucose sensing shapes T cell immune responses. Glucose transporter 1 (Glut1) transports glucose into T cells to fuel glycolysis. The generation of effector T cells, including TH1, TH2, TH17, and TFH cells, is dependent on Glut1 expression and glycolytic metabolism. While Glut1 expression and glycolysis promote Treg cell proliferation, they impair Treg cell lineage stability and suppressive function. Thus, Glut1 expression provides a key mechanism to shape T cell responses. Glycolysis also promotes the formation of effector memory CD8+ T cells (TEM). TCR and CD28 co-stimulation potently induces Glut1 expression, while the translocation of Glut1 from the cytoplasm to T cell surface is induced by various stimuli, including TCR-CD28, IL-7, and insulin.
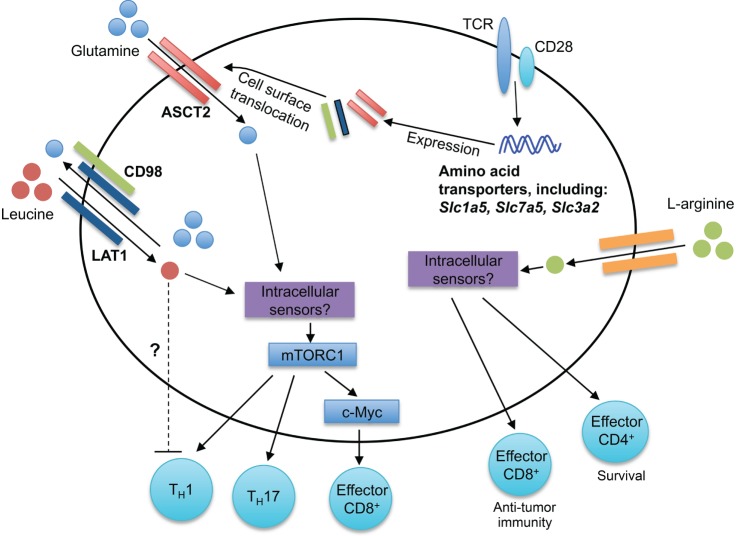
Amino acid sensing modulates T cell responses. Antigen-driven activation of T cells through TCRs upregulates expression of many amino acid transporters, including the leucine and glutamine transporters LAT1, ASCT2, and CD98. LAT1 associates with CD98, forming a bidirectional transporter for leucine and glutamine. The intracellular sensors of leucine and glutamine in T cells remain unknown. mTORC1 is activated downstream of intracellular amino acids, leading to the regulation of CD4+ T cell differentiation (TH1 and TH17) and CD8+ T cell effector responses. Elevated intracellular l -arginine levels promote effector CD8+ T cell anti-tumor immunity and effector CD4+ T cell IFN-γ production and survival. Although the intracellular mediators critical for arginine sensing are unclear, three potential sensors are BAZ1B, PSIP1, and TSN.
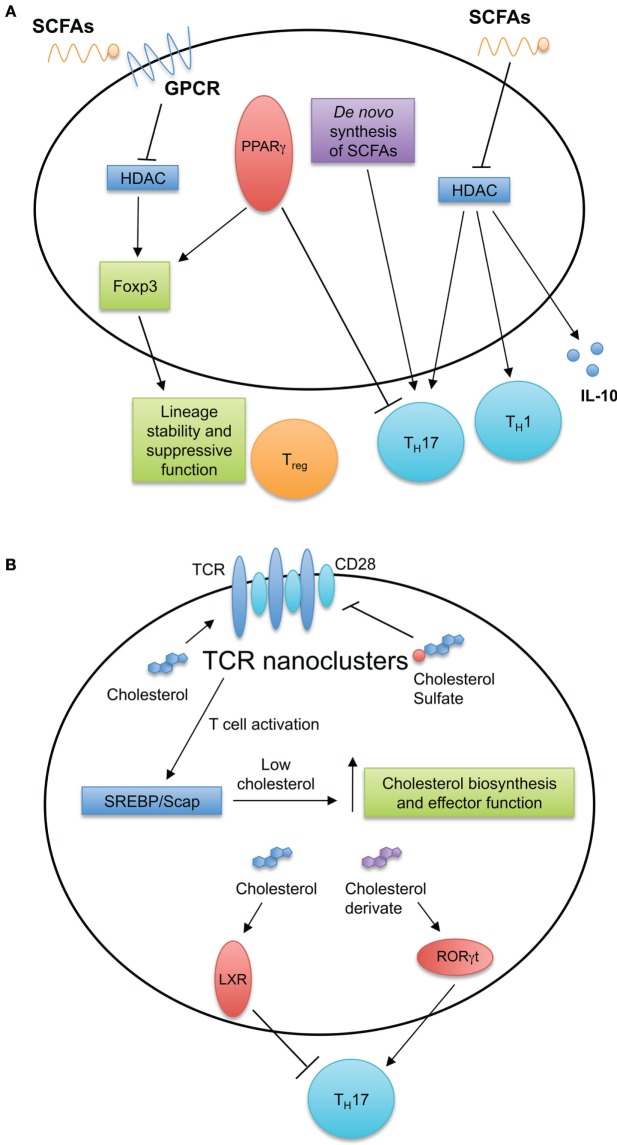
Fatty acid (FA) and cholesterol sensing in T cell immune responses. (A) FA metabolism and sensing serve multiple roles in T cell biology. Short-chain fatty acids (SCFAs) diffuse passively or are transported into cells by G-protein-coupled receptors. SCFAs can inhibit histone deacetylases and induce TH1 and TH17 cell differentiation and IL-10 production. De novo FA synthesis is required for TH17 generation. Intracellular FAs are recognized by peroxisome proliferator-activated receptors (PPARs). Expression of PPARγ is important for visceral adipose tissue Treg cell accumulation and function, while antagonizing PPARγ by chemical drug promotes TH17 development. (B) Cholesterol and cholesterol sulfates regulate T cell receptor (TCR) nanoclustering and signaling within the T cell. After stimulation of TCRs, T cells upregulate the cholesterol biosynthesis pathway through the SREBP/SCAP axis. This increase in cholesterol biosynthesis is important for acquiring T cell effector function. Additionally, intracellular sterols serve as ligands for intracellular receptors and transcription factors, such as liver X receptor (LXR) and RORγt to modulate TH17 development.
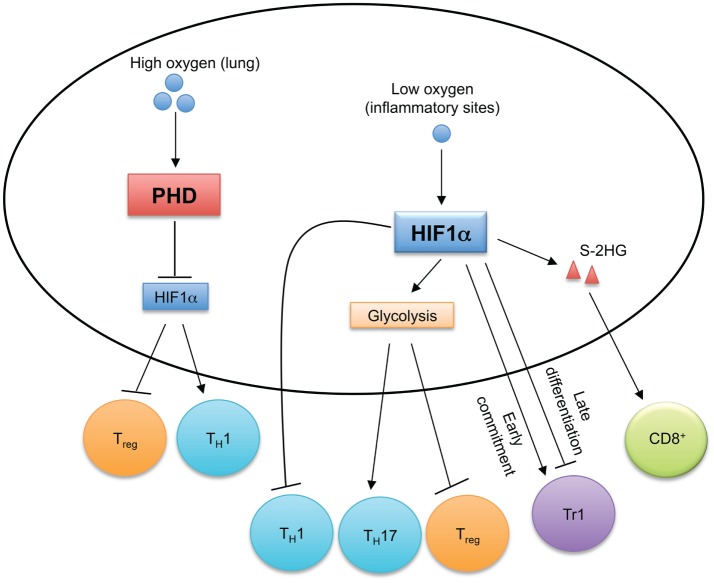
Oxygen sensing orchestrates T cell immune responses. In oxygen-replete environments, such as in the lung, prolyl-hydroxylase domain (PHD) proteins sense high levels of cellular oxygen. Activated PHD proteins establish immune tolerance in the lung by inducing hypoxia-inducible factor 1α (HIF1α) degradation to promote Treg cell and inhibit TH1 cell differentiation. Hypoxia induces HIF1α expression, which regulates the reciprocal generation of TH17 and Treg cells and antagonizes TH1 cell differentiation. HIF1α also temporally regulates Tr1 cell differentiation. HIF1α-dependent upregulation of S-2-hydroxyglutarate (S-2HG) enhances anti-tumor CD8+ T cell responses.
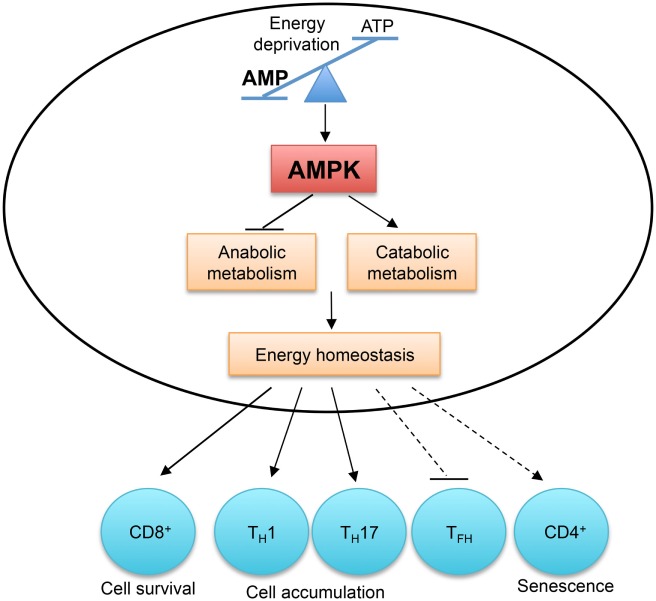
AMP-activated protein kinase (AMPK) and energy sensing dictate T cell immunity. AMPK is activated by the increased AMP to ATP ratio, which indicates energy deprivation. AMPK functions as a master regulator of cellular metabolism to preserve cellular energy homeostasis. AMPK activity is required for CD8+ T cell survival during infection and in tumor microenvironment. The accumulation of TH1 and TH17 cells in the colon is also dependent on AMPK activity. Aberrant AMPK activation is linked to impaired TFH cell generation and CD4+ T cell senescence.
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources