

Lipid-dependent conformational dynamics underlie the functional versatility of T-cell receptor
- PMID: 28337984
- PMCID: PMC5385618
- DOI: 10.1038/cr.2017.42
Lipid-dependent conformational dynamics underlie the functional versatility of T-cell receptor
Abstract
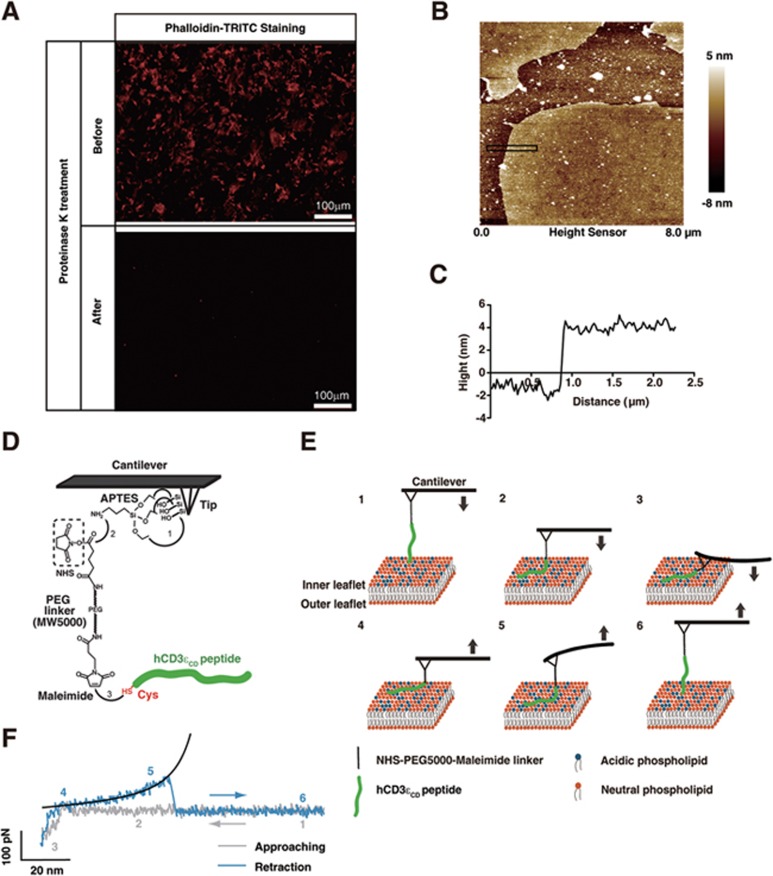
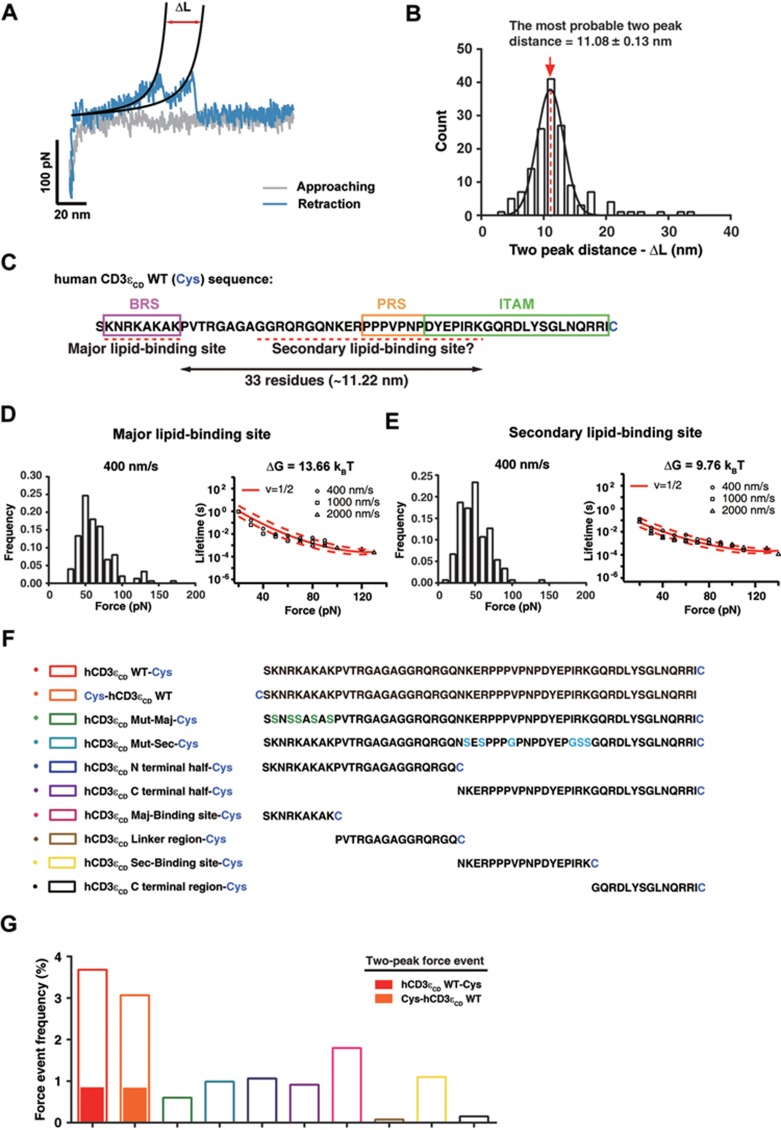
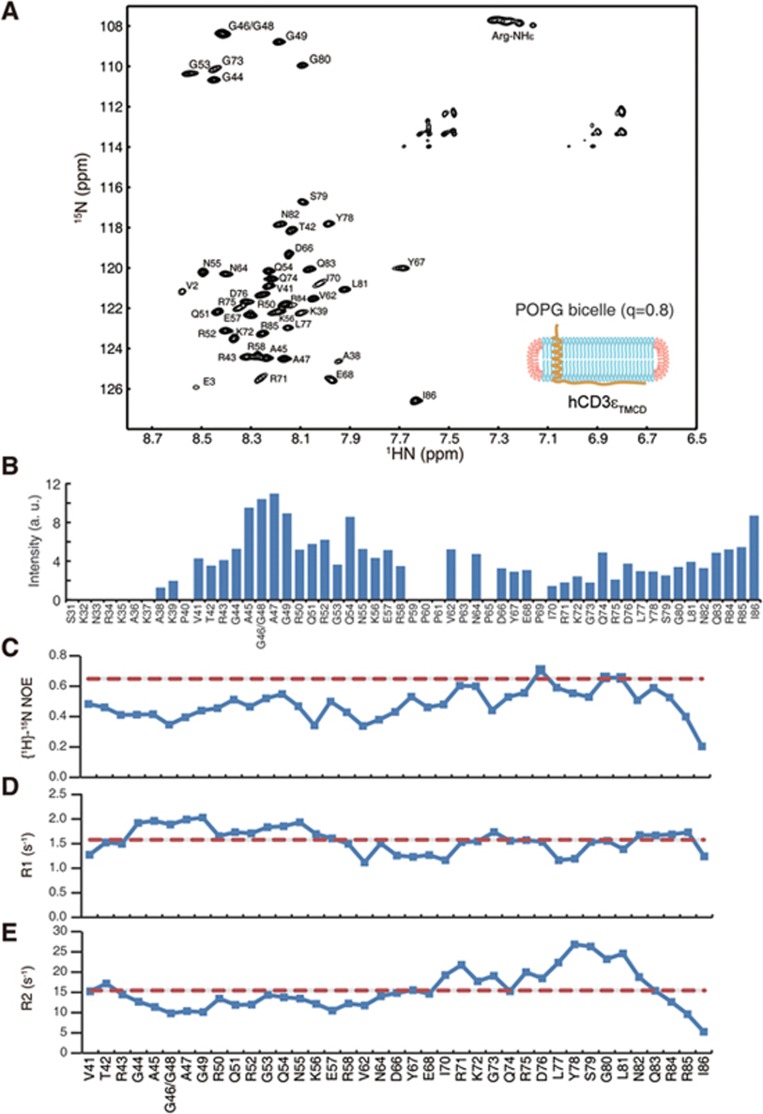
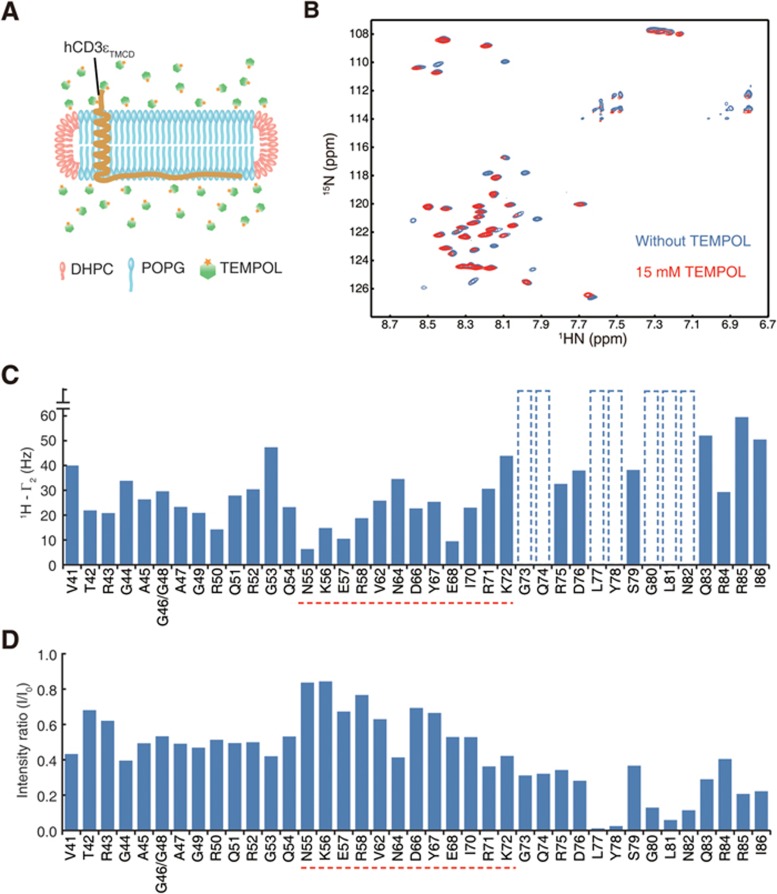
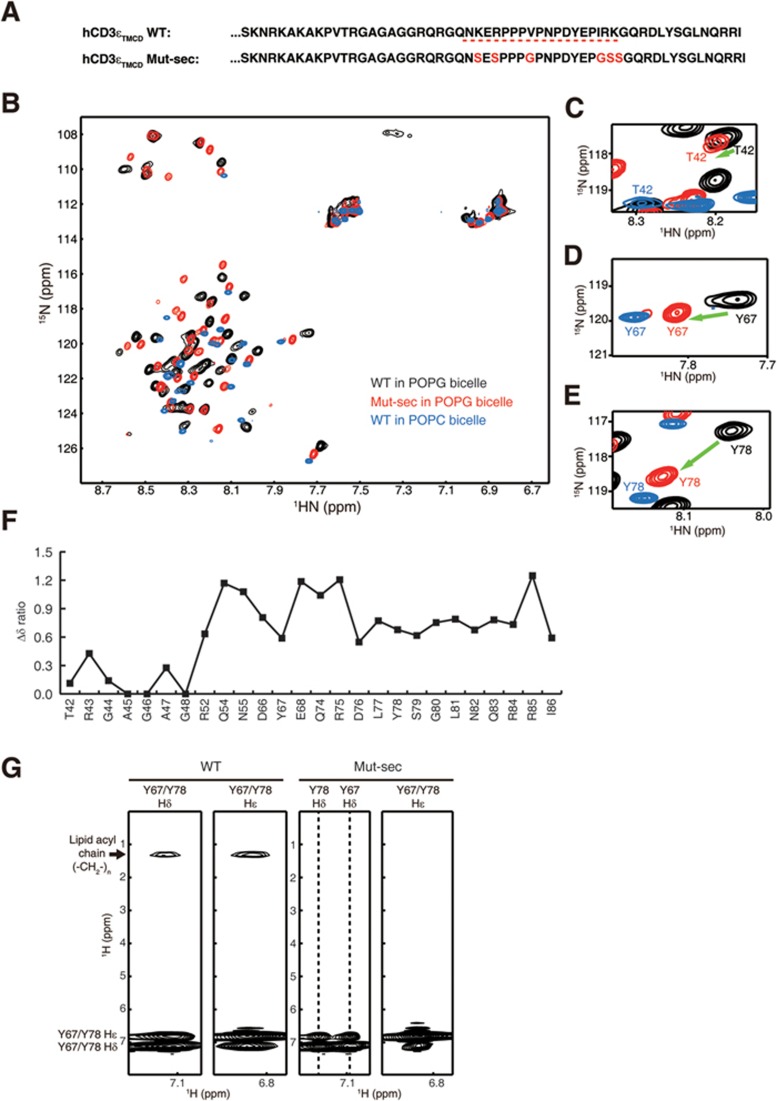
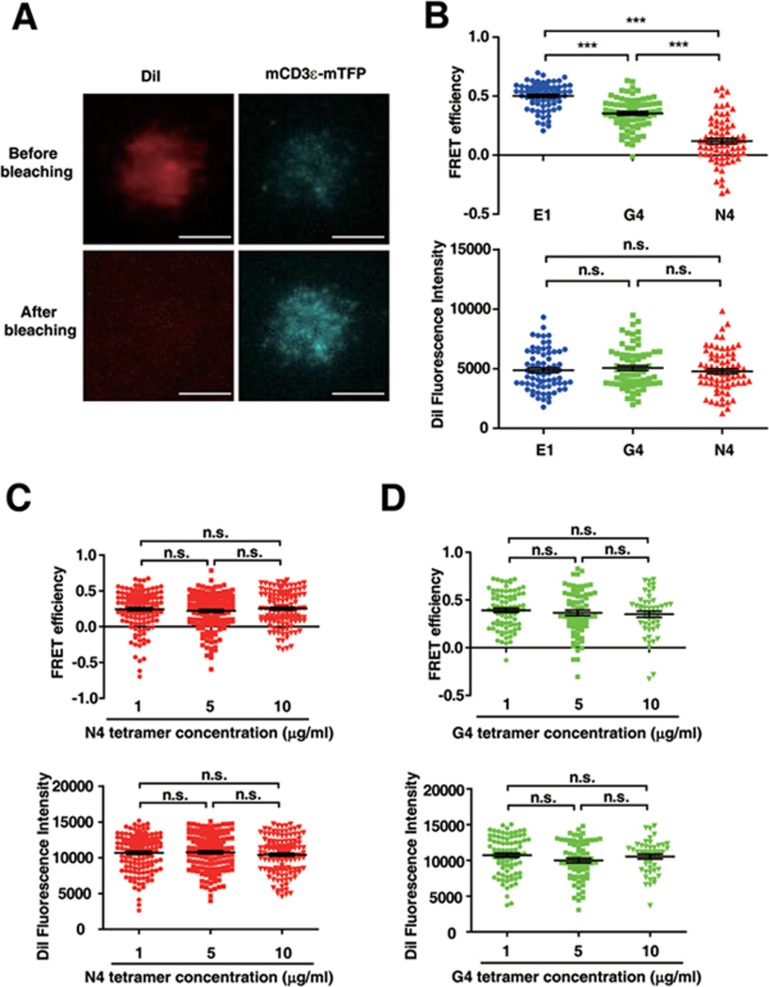
T-cell receptor-CD3 complex (TCR) is a versatile signaling machine that can initiate antigen-specific immune responses based on various biochemical changes of CD3 cytoplasmic domains, but the underlying structural basis remains elusive. Here we developed biophysical approaches to study the conformational dynamics of CD3ε cytoplasmic domain (CD3εCD). At the single-molecule level, we found that CD3εCD could have multiple conformational states with different openness of three functional motifs, i.e., ITAM, BRS and PRS. These conformations were generated because different regions of CD3εCD had heterogeneous lipid-binding properties and therefore had heterogeneous dynamics. Live-cell imaging experiments demonstrated that different antigen stimulations could stabilize CD3εCD at different conformations. Lipid-dependent conformational dynamics thus provide structural basis for the versatile signaling property of TCR.
Figures

References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
