

Septin 9_i2 is downregulated in tumors, impairs cancer cell migration and alters subnuclear actin filaments
- PMID: 28338090
- PMCID: PMC5364497
- DOI: 10.1038/srep44976
Septin 9_i2 is downregulated in tumors, impairs cancer cell migration and alters subnuclear actin filaments
Abstract
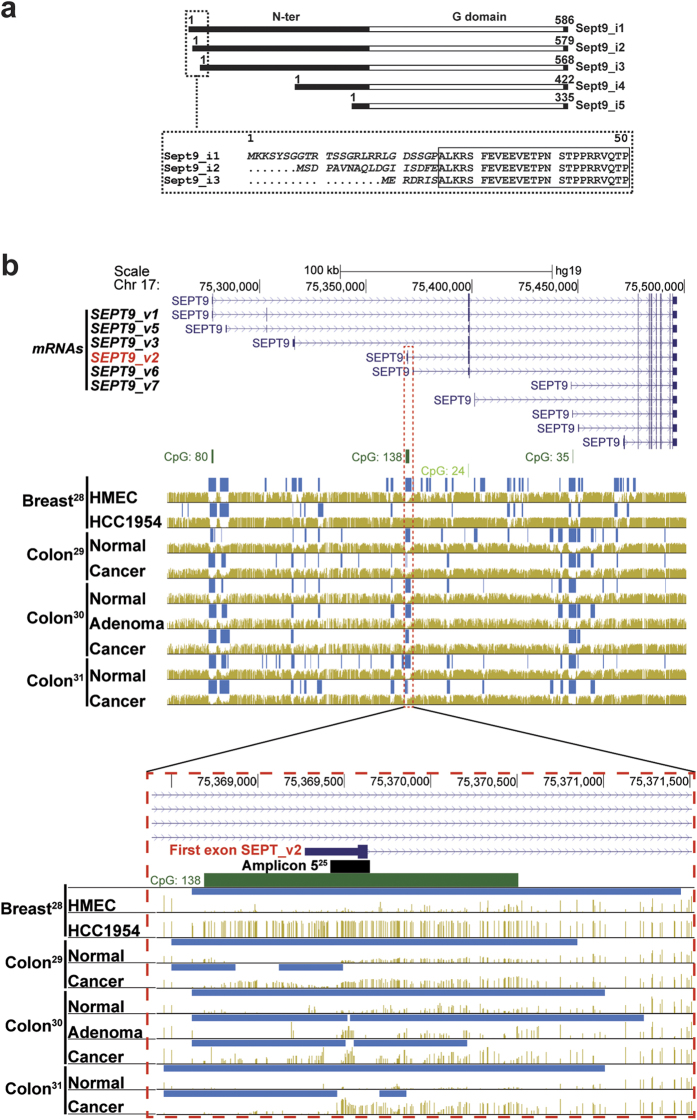
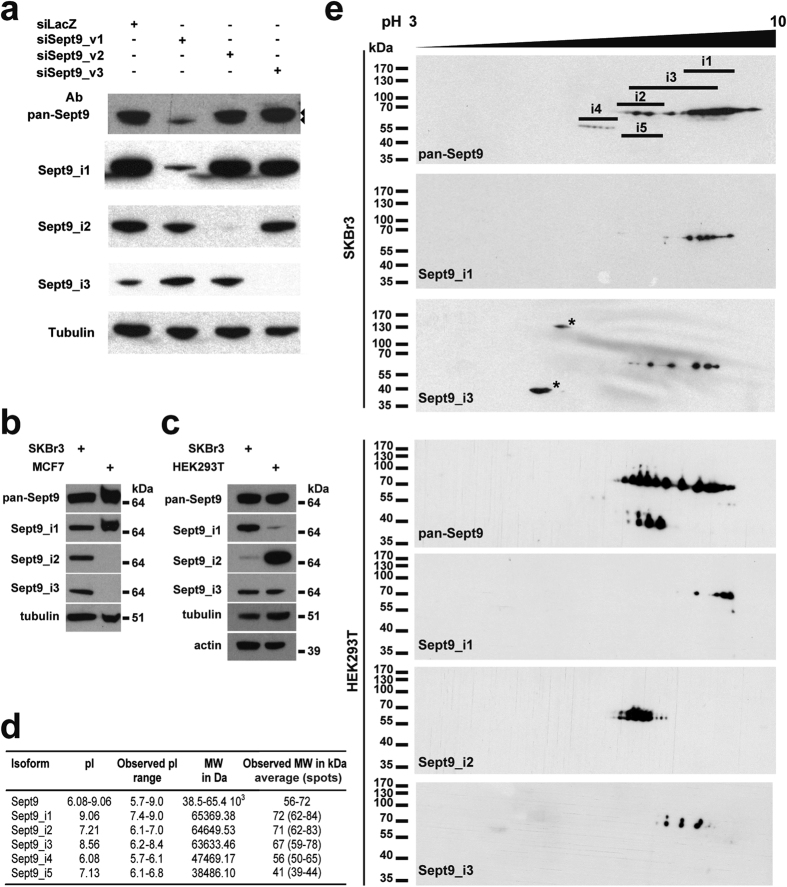
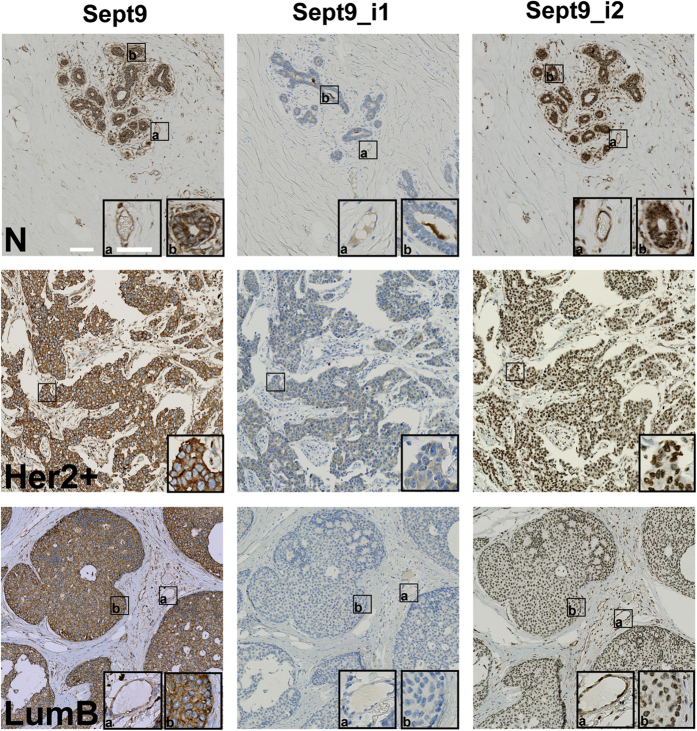
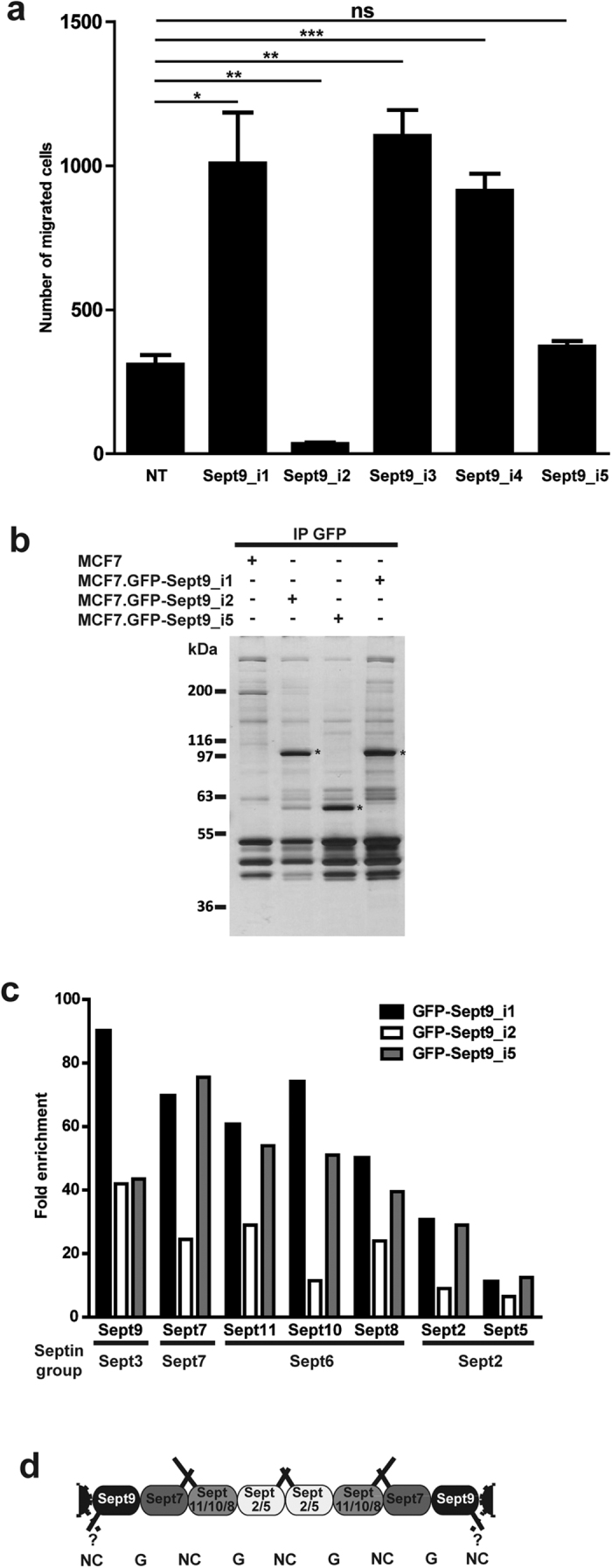
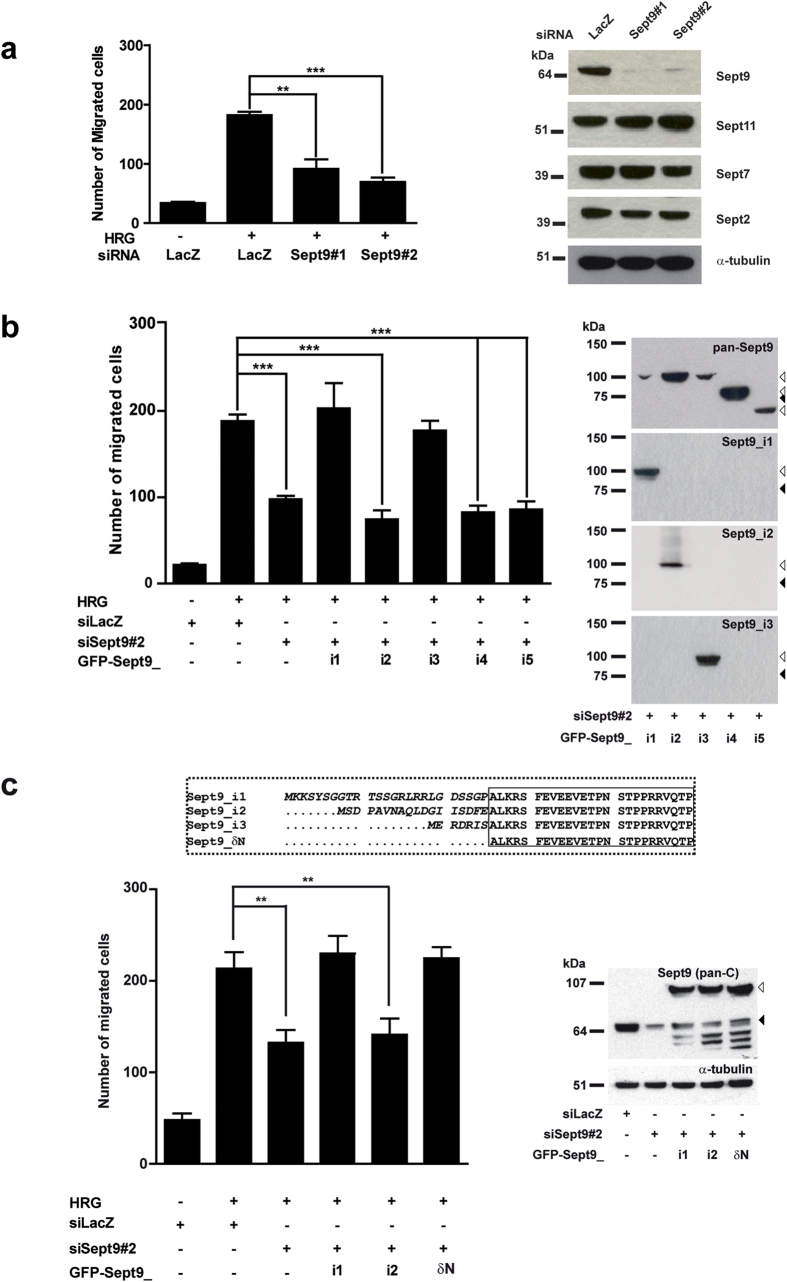
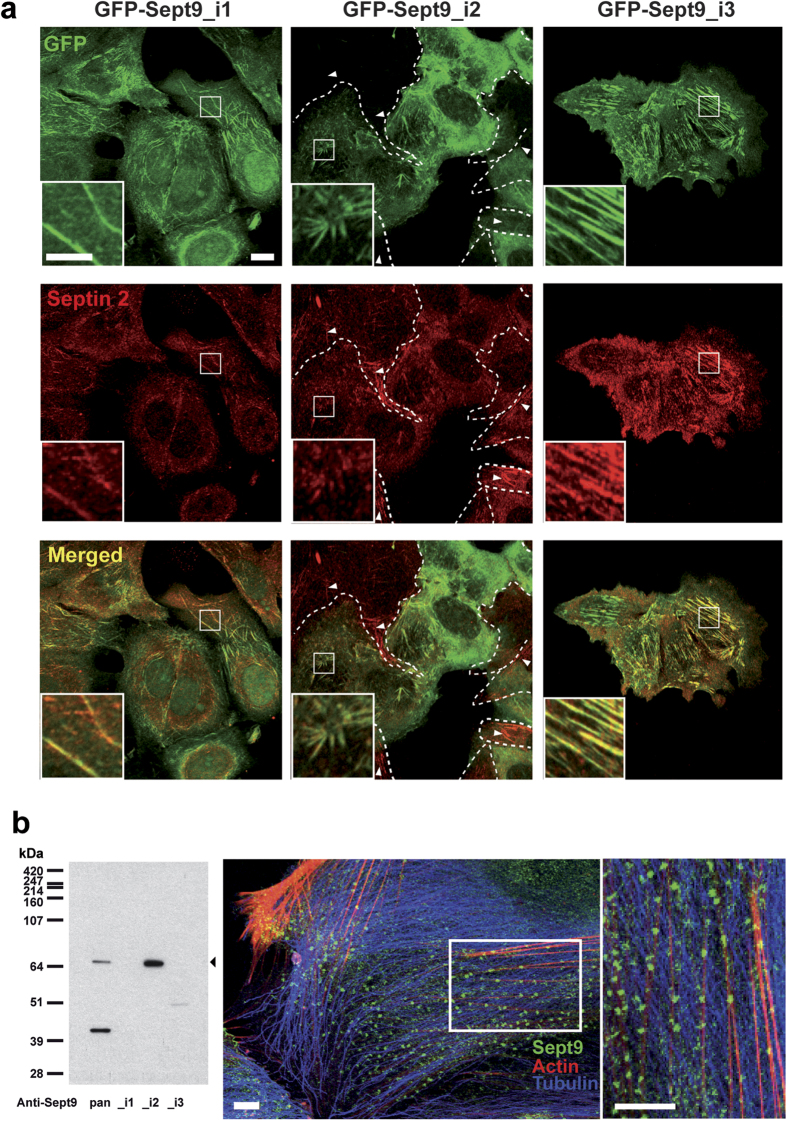
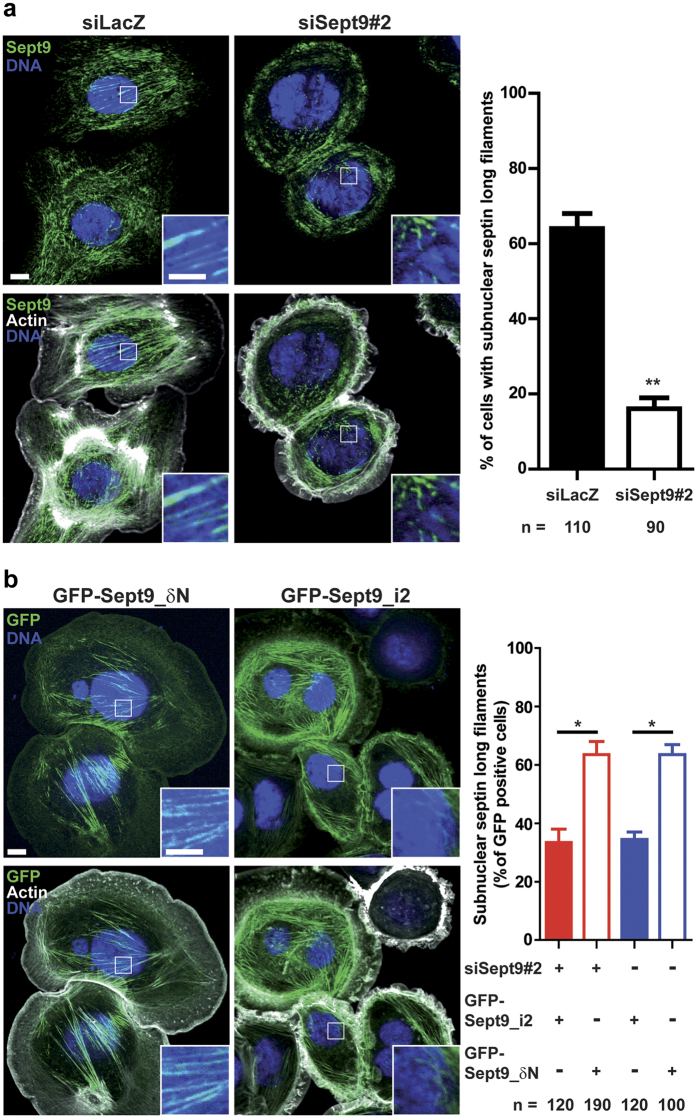
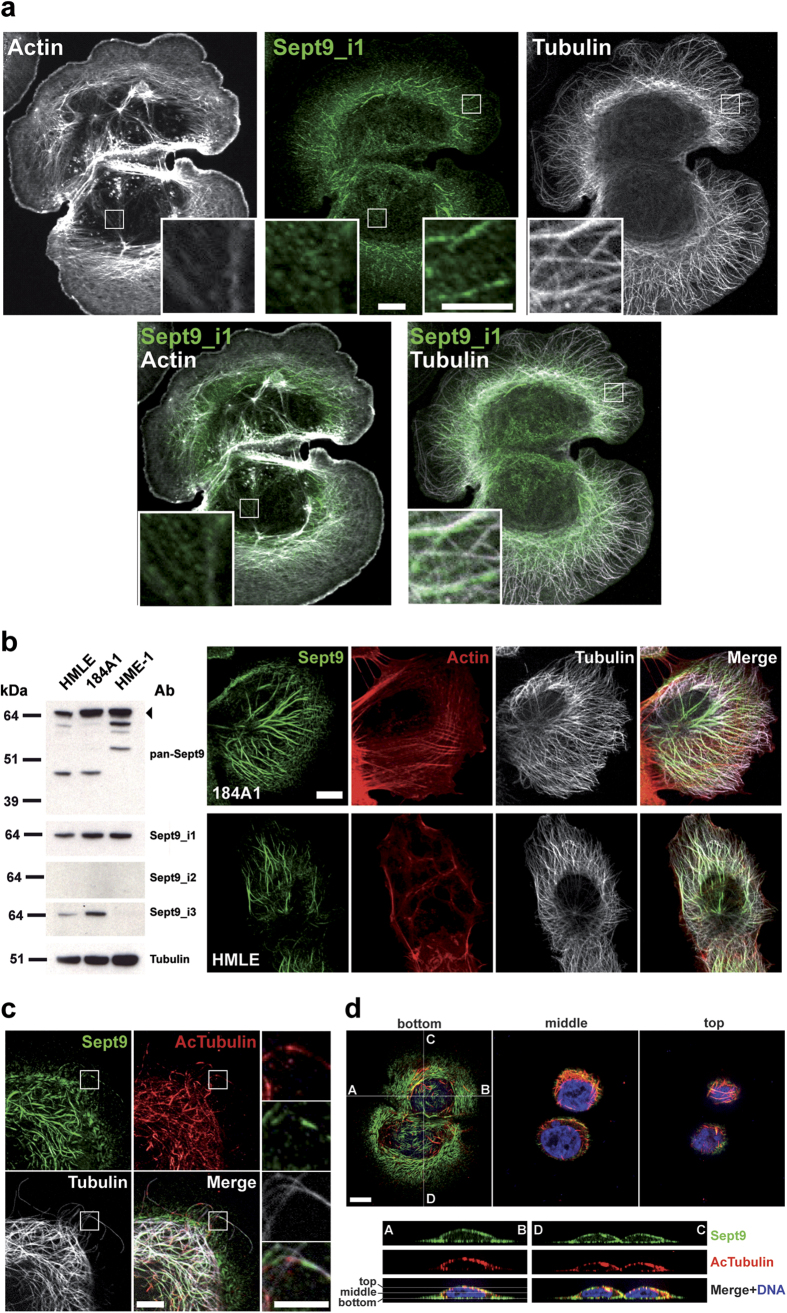
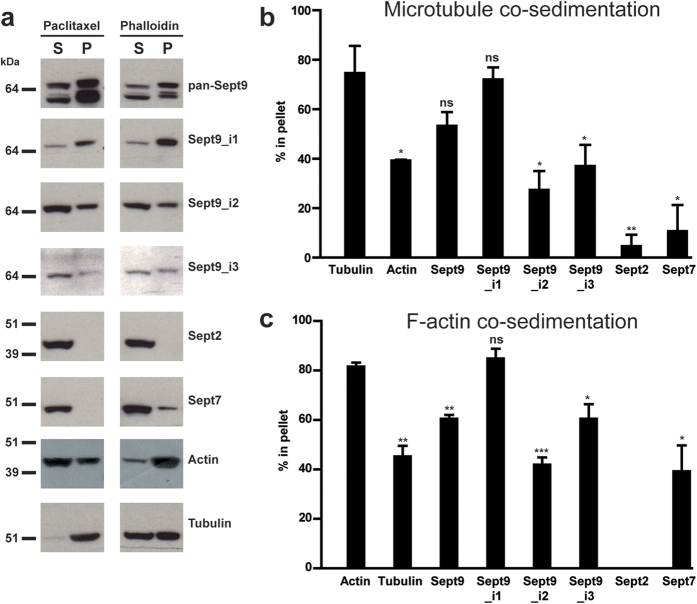
Functions of septin cytoskeletal polymers in tumorigenesis are still poorly defined. Their role in the regulation of cytokinesis and cell migration were proposed to contribute to cancer associated aneuploidy and metastasis. Overexpression of Septin 9 (Sept9) promotes migration of cancer cell lines. SEPT9 mRNA and protein expression is increased in breast tumors compared to normal and peritumoral tissues and amplification of SEPT9 gene was positively correlated with breast tumor progression. However, the existence of multiple isoforms of Sept9 is a confounding factor in the analysis of Sept9 functions. In the present study, we analyze the protein expression of Sept9_i2, an uncharacterized isoform, in breast cancer cell lines and tumors and describe its specific impact on cancer cell migration and Sept9 cytoskeletal distribution. Collectively, our results showed that, contrary to Sept9_i1, Sept9_i2 did not support cancer cell migration, and induced a loss of subnuclear actin filaments. These effects were dependent on Sept9_i2 specific N-terminal sequence. Sept9_i2 was strongly down-regulated in breast tumors compared to normal mammary tissues. Thus our data indicate that Sept9_i2 is a negative regulator of breast tumorigenesis. We propose that Sept9 tumorigenic properties depend on the balance between Sept9_i1 and Sept9_i2 expression levels.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Mostowy S. & Cossart P. Septins: the fourth component of the cytoskeleton. Nat. Rev. Mol. Cell Biol. 13, 183–194 (2012). - PubMed
-
- Fung K. Y., Dai L. & Trimble W. S. Cell and molecular biology of septins. Int. Rev. Cell. Mol. Biol. 310, 289–339 (2014). - PubMed
-
- Kinoshita M., Field C. M., Coughlin M. L., Straight A. F. & Mitchison T. J. Self- and actin-templated assembly of Mammalian septins. Dev. Cell 3, 791–802 (2002). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
