

Intermediate filament reorganization dynamically influences cancer cell alignment and migration
- PMID: 28338091
- PMCID: PMC5364536
- DOI: 10.1038/srep45152
Intermediate filament reorganization dynamically influences cancer cell alignment and migration
Abstract
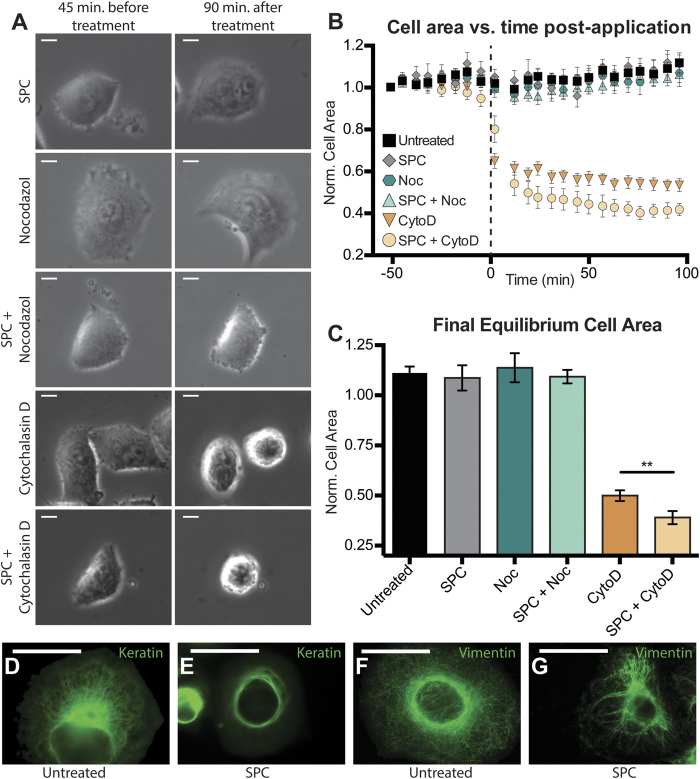
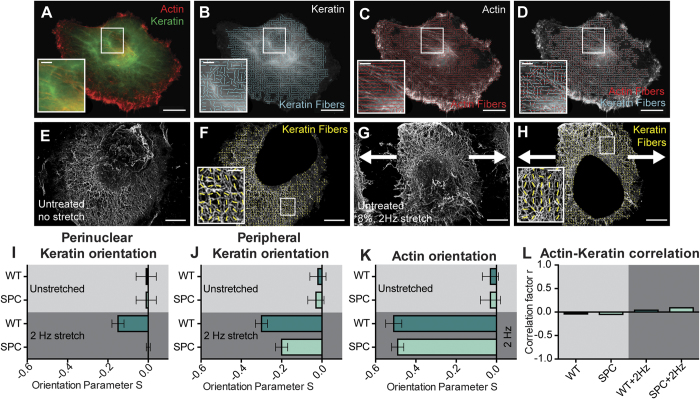
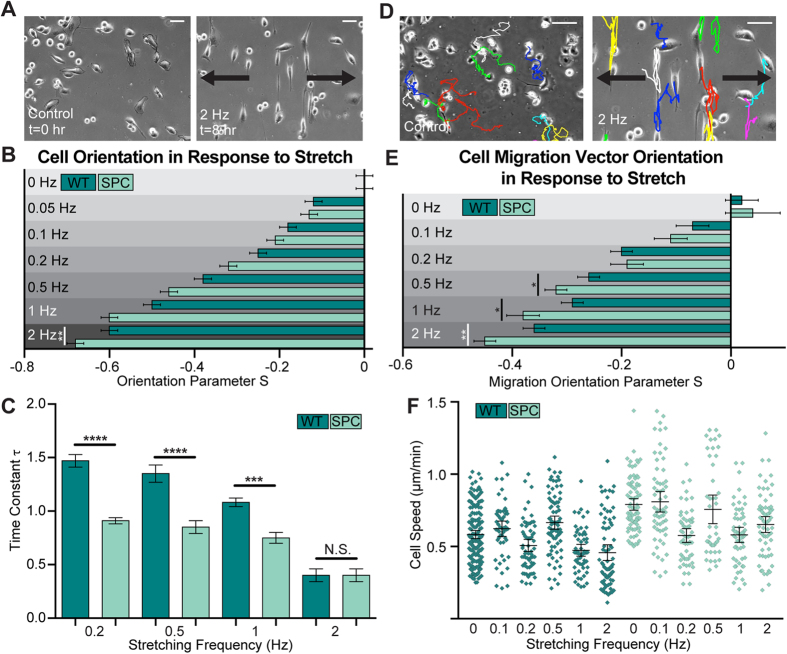
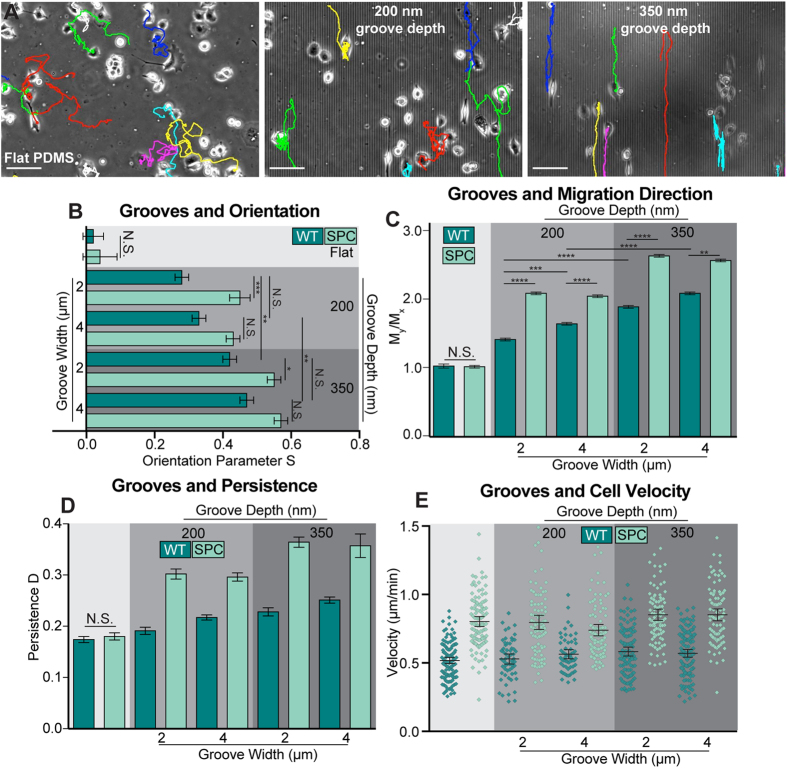
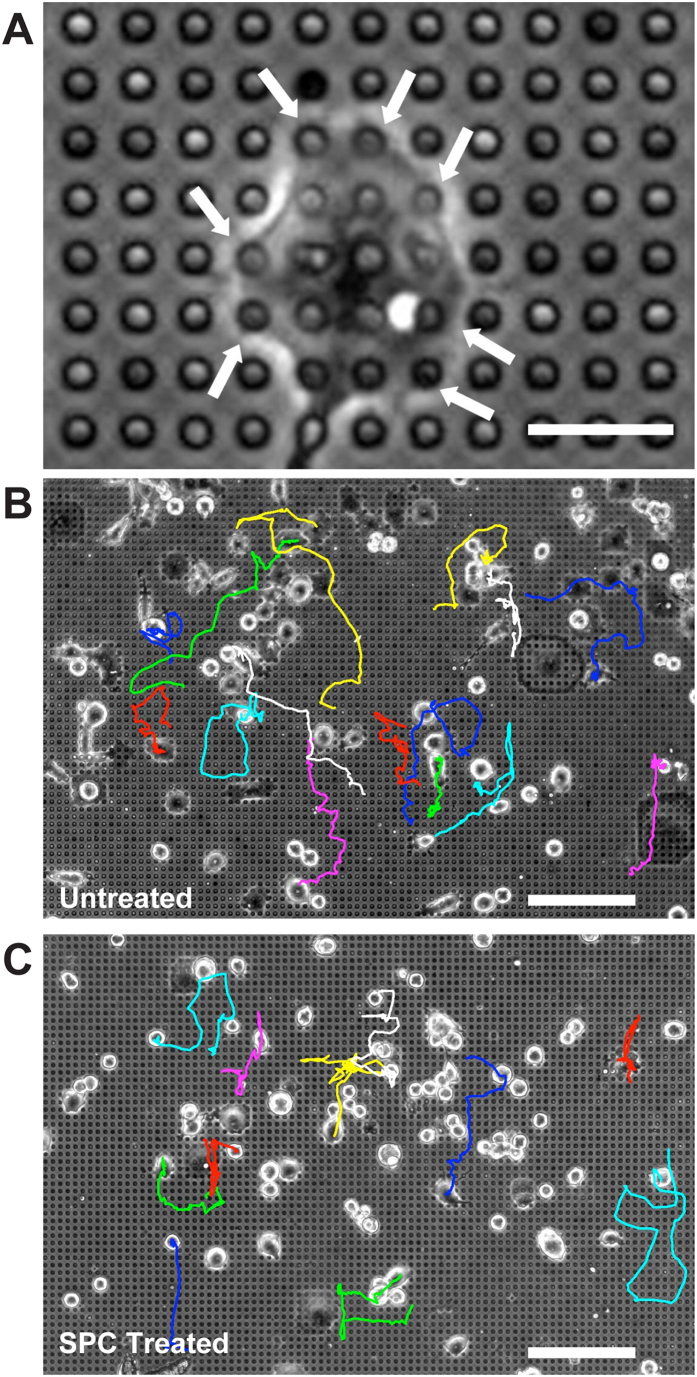
The interactions between a cancer cell and its extracellular matrix (ECM) have been the focus of an increasing amount of investigation. The role of the intermediate filament keratin in cancer has also been coming into focus of late, but more research is needed to understand how this piece fits in the puzzle of cytoskeleton-mediated invasion and metastasis. In Panc-1 invasive pancreatic cancer cells, keratin phosphorylation in conjunction with actin inhibition was found to be sufficient to reduce cell area below either treatment alone. We then analyzed intersecting keratin and actin fibers in the cytoskeleton of cyclically stretched cells and found no directional correlation. The role of keratin organization in Panc-1 cellular morphological adaptation and directed migration was then analyzed by culturing cells on cyclically stretched polydimethylsiloxane (PDMS) substrates, nanoscale grates, and rigid pillars. In general, the reorganization of the keratin cytoskeleton allows the cell to become more 'mobile'- exhibiting faster and more directed migration and orientation in response to external stimuli. By combining keratin network perturbation with a variety of physical ECM signals, we demonstrate the interconnected nature of the architecture inside the cell and the scaffolding outside of it, and highlight the key elements facilitating cancer cell-ECM interactions.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
