

Modular Organization of Residue-Level Contacts Shapes the Selection Pressure on Individual Amino Acid Sites of Ribosomal Proteins
- PMID: 28338825
- PMCID: PMC5388290
- DOI: 10.1093/gbe/evx036
Modular Organization of Residue-Level Contacts Shapes the Selection Pressure on Individual Amino Acid Sites of Ribosomal Proteins
Abstract
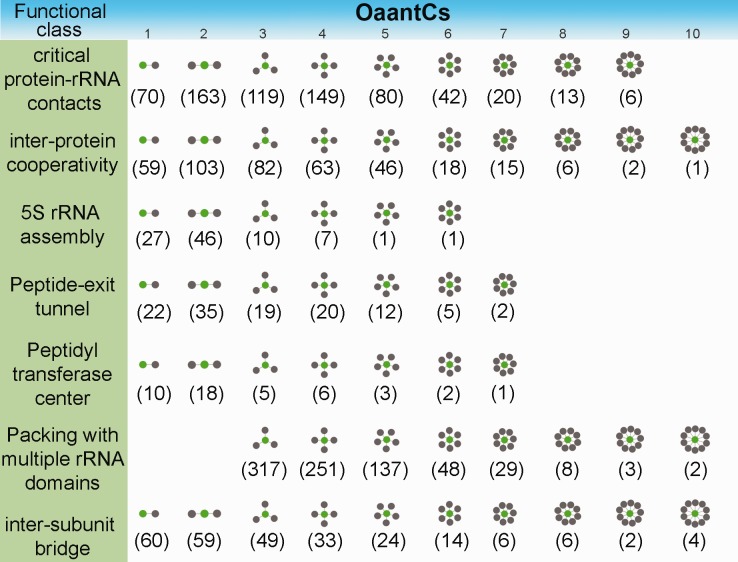
Understanding the molecular evolution of macromolecular complexes in the light of their structure, assembly, and stability is of central importance. Here, we address how the modular organization of native molecular contacts shapes the selection pressure on individual residue sites of ribosomal complexes. The bacterial ribosomal complex is represented as a residue contact network where nodes represent amino acid/nucleotide residues and edges represent their van der Waals interactions. We find statistically overrepresented native amino acid-nucleotide contacts (OaantC, one amino acid contacts one or multiple nucleotides, internucleotide contacts are disregarded). Contact number is defined as the number of nucleotides contacted. Involvement of individual amino acids in OaantCs with smaller contact numbers is more random, whereas only a few amino acids significantly contribute to OaantCs with higher contact numbers. An investigation of structure, stability, and assembly of bacterial ribosome depicts the involvement of these OaantCs in diverse biophysical interactions stabilizing the complex, including high-affinity protein-RNA contacts, interprotein cooperativity, intersubunit bridge, packing of multiple ribosomal RNA domains, etc. Amino acid-nucleotide constituents of OaantCs with higher contact numbers are generally associated with significantly slower substitution rates compared with that of OaantCs with smaller contact numbers. This evolutionary rate heterogeneity emerges from the strong purifying selection pressure that conserves the respective amino acid physicochemical properties relevant to the stabilizing interaction with OaantC nucleotides. An analysis of relative molecular orientations of OaantC residues and their interaction energetics provides the biophysical ground of purifying selection conserving OaantC amino acid physicochemical properties.
Keywords: contact network; network motif; protein–RNA interaction; purifying selection; ribosome; stability.
© The Author(s) 2017. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
