

RNA Editing During Sexual Development Occurs in Distantly Related Filamentous Ascomycetes
- PMID: 28338982
- PMCID: PMC5381528
- DOI: 10.1093/gbe/evx052
RNA Editing During Sexual Development Occurs in Distantly Related Filamentous Ascomycetes
Abstract
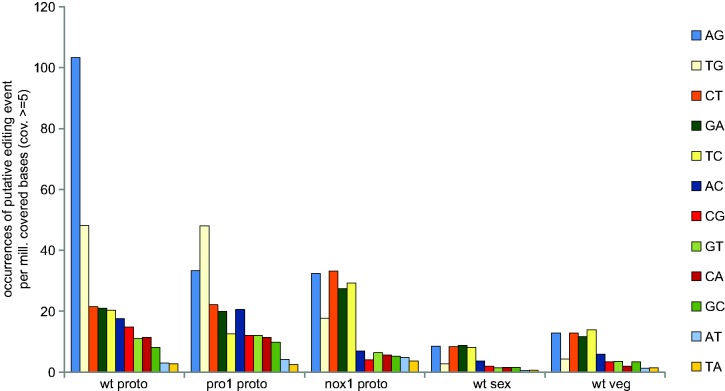
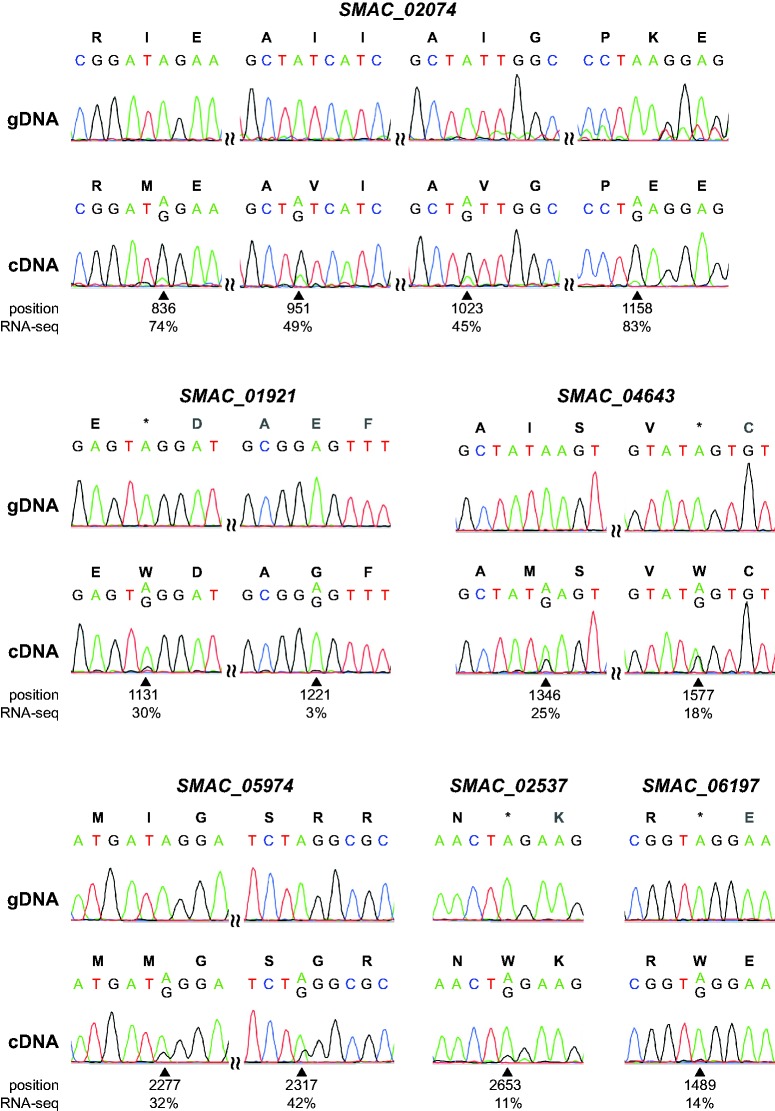
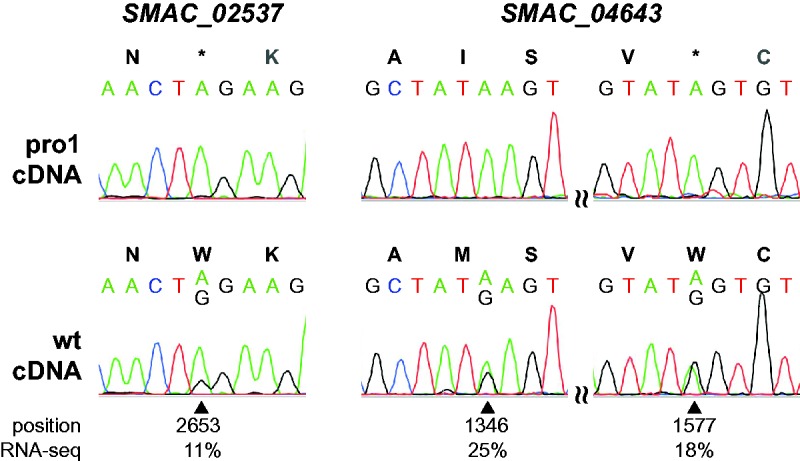
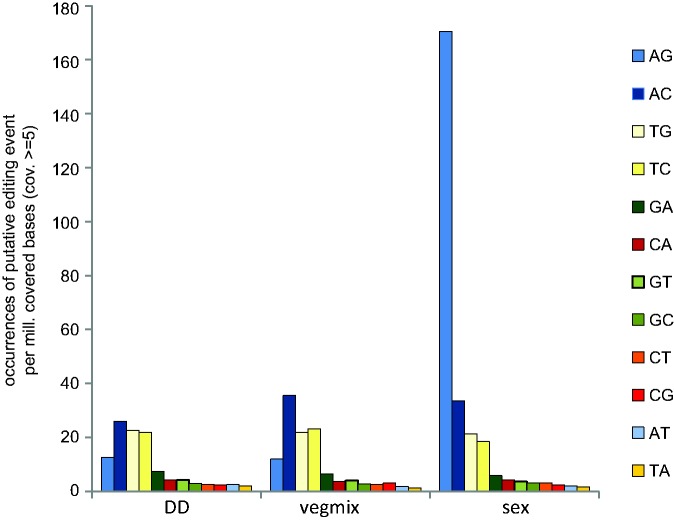
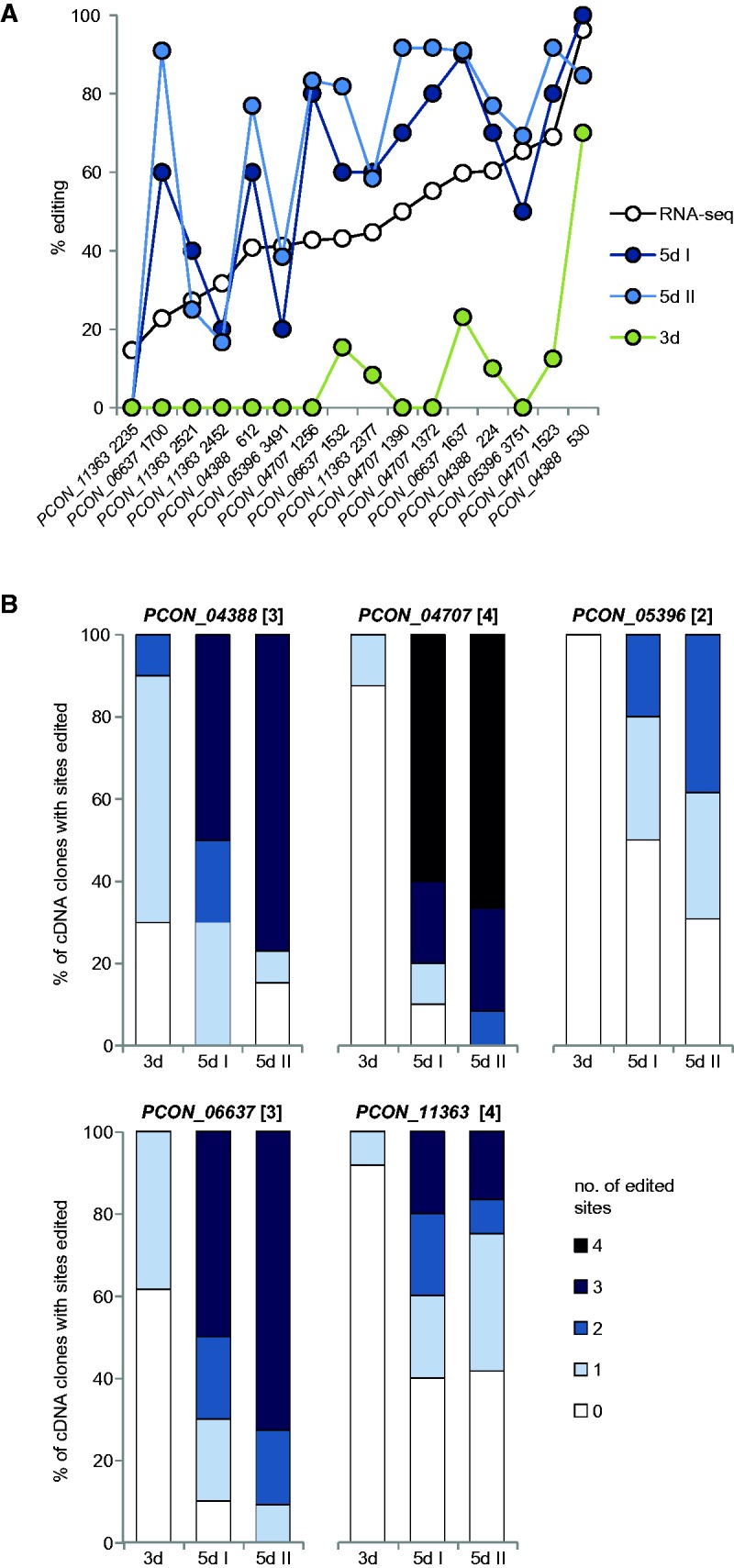
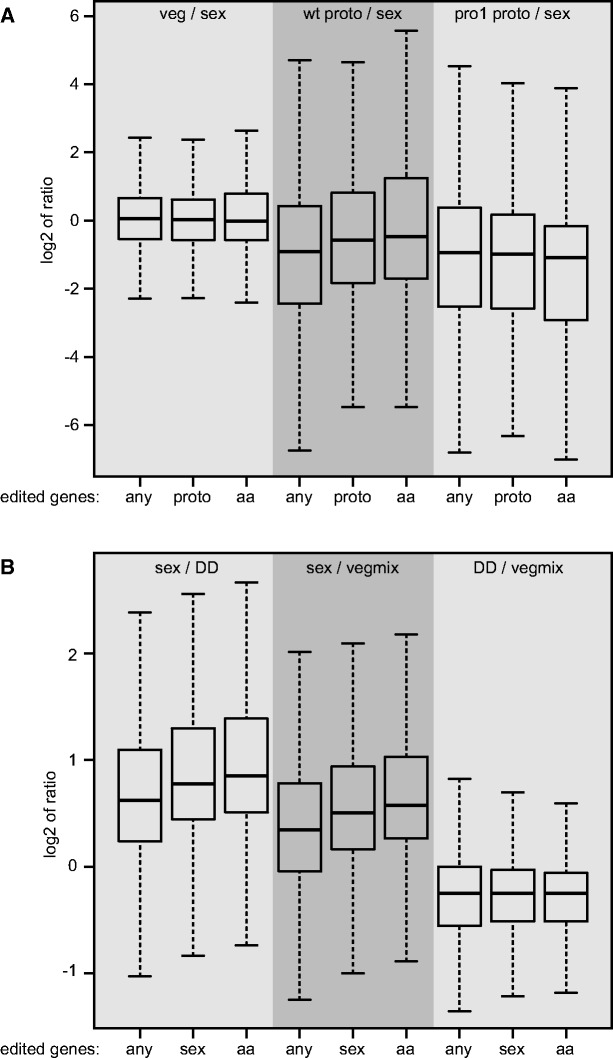
RNA editing is a post-transcriptional process that modifies RNA molecules leading to transcript sequences that differ from their template DNA. A-to-I editing was found to be widely distributed in nuclear transcripts of metazoa, but was detected in fungi only recently in a study of the filamentous ascomycete Fusarium graminearum that revealed extensive A-to-I editing of mRNAs in sexual structures (fruiting bodies). Here, we searched for putative RNA editing events in RNA-seq data from Sordaria macrospora and Pyronema confluens, two distantly related filamentous ascomycetes, and in data from the Taphrinomycete Schizosaccharomyces pombe. Like F. graminearum, S. macrospora is a member of the Sordariomycetes, whereas P. confluens belongs to the early-diverging group of Pezizomycetes. We found extensive A-to-I editing in RNA-seq data from sexual mycelium from both filamentous ascomycetes, but not in vegetative structures. A-to-I editing was not detected in different stages of meiosis of S. pombe. A comparison of A-to-I editing in S. macrospora with F. graminearum and P. confluens, respectively, revealed little conservation of individual editing sites. An analysis of RNA-seq data from two sterile developmental mutants of S. macrospora showed that A-to-I editing is strongly reduced in these strains. Sequencing of cDNA fragments containing more than one editing site from P. confluens showed that at the beginning of sexual development, transcripts were incompletely edited or unedited, whereas in later stages transcripts were more extensively edited. Taken together, these data suggest that A-to-I RNA editing is an evolutionary conserved feature during fruiting body development in filamentous ascomycetes.
Keywords: A-to-I RNA editing; Pyronema confluens; Sordaria macrospora; fruiting body; protoperithecia; sexual development.
© The Author(s) 2017. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
