

Z-Guggulsterone Produces Antidepressant-Like Effects in Mice through Activation of the BDNF Signaling Pathway
- PMID: 28339691
- PMCID: PMC5458345
- DOI: 10.1093/ijnp/pyx009
Z-Guggulsterone Produces Antidepressant-Like Effects in Mice through Activation of the BDNF Signaling Pathway
Abstract
Background: Z-guggulsterone, an active compound extracted from the gum resin of the tree Commiphora mukul, has been shown to improve animal memory deficits via activating the brain-derived neurotrophic factor signaling pathway. Here, we investigated the antidepressant-like effect of Z-guggulsterone in a chronic unpredictable stress mouse model of depression.
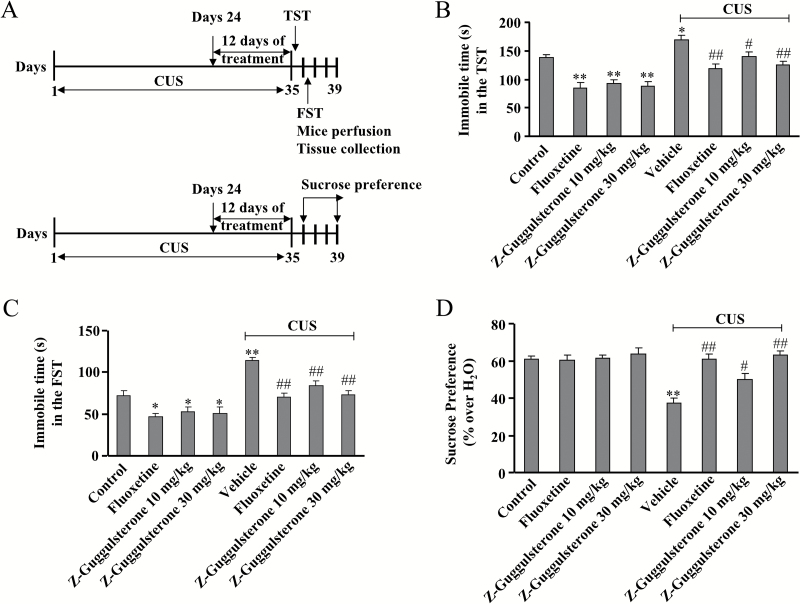
Methods: The effects of Z-guggulsterone were assessed in mice with the tail suspension test and forced swimming test. Z-guggulsterone was also investigated in the chronic unpredictable stress model of depression with fluoxetine as the positive control. Changes in hippocampal neurogenesis as well as the brain-derived neurotrophic factor signaling pathway after chronic unpredictable stress/Z-guggulsterone treatment were investigated. The tryptophan hydroxylase inhibitor and the tyrosine kinase B inhibitor were also used to explore the antidepressant-like mechanisms of Z-guggulsterone.
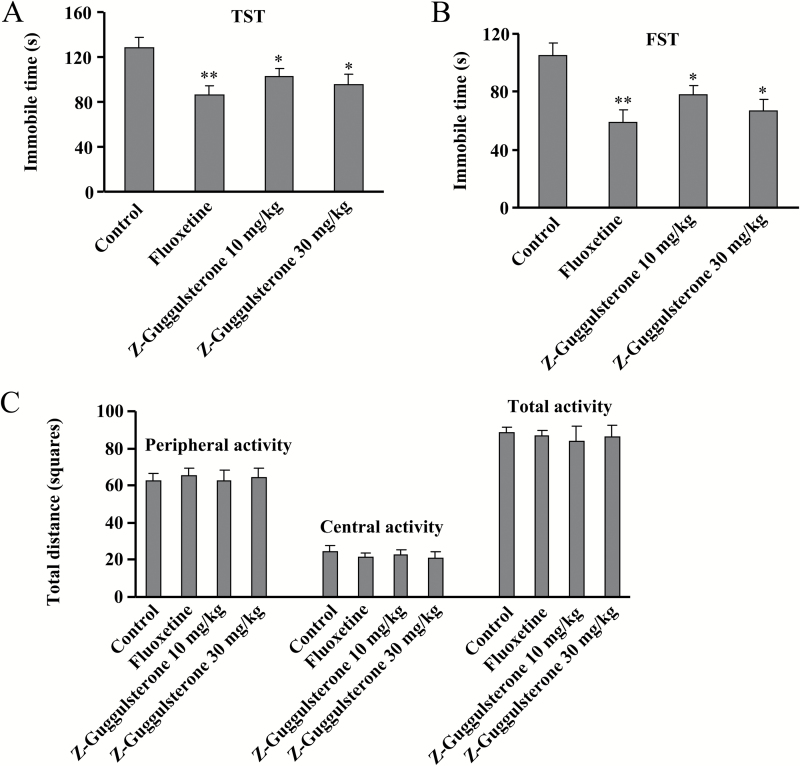
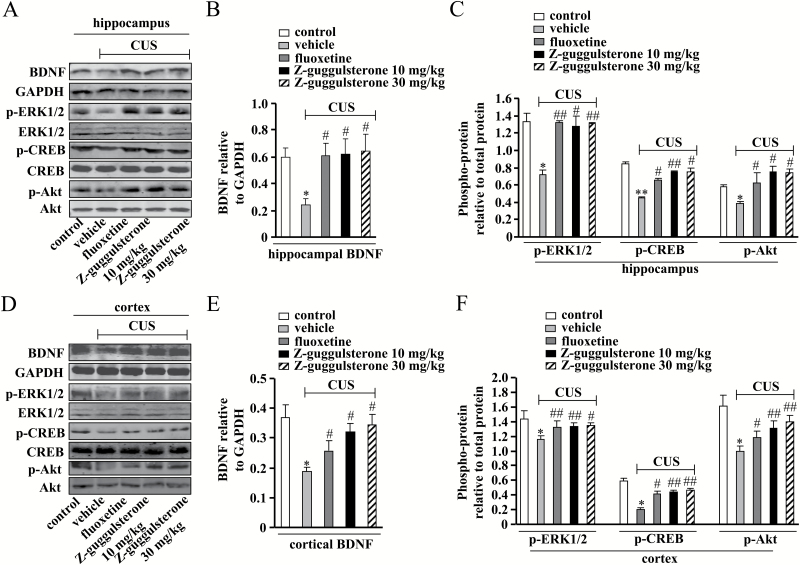
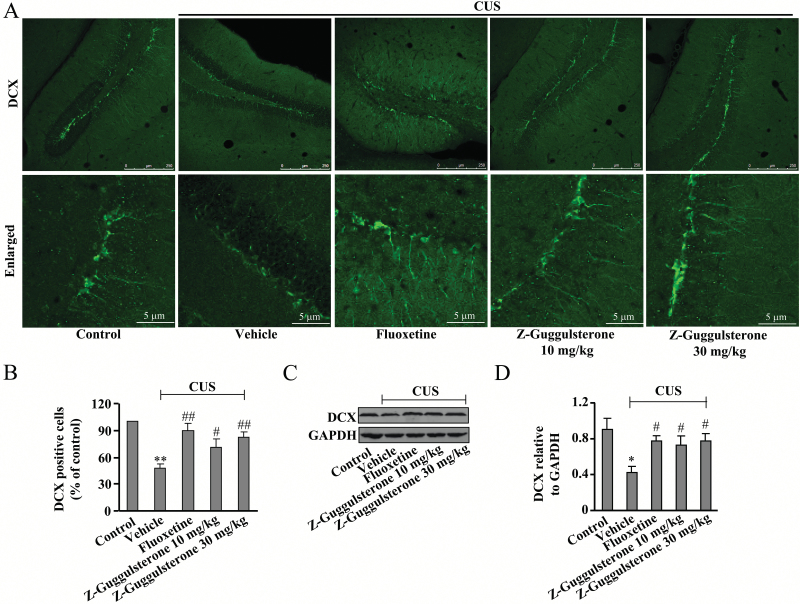
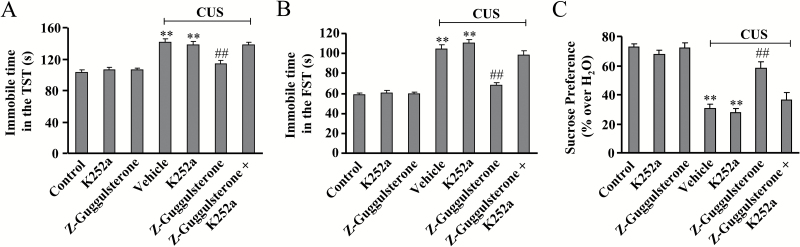
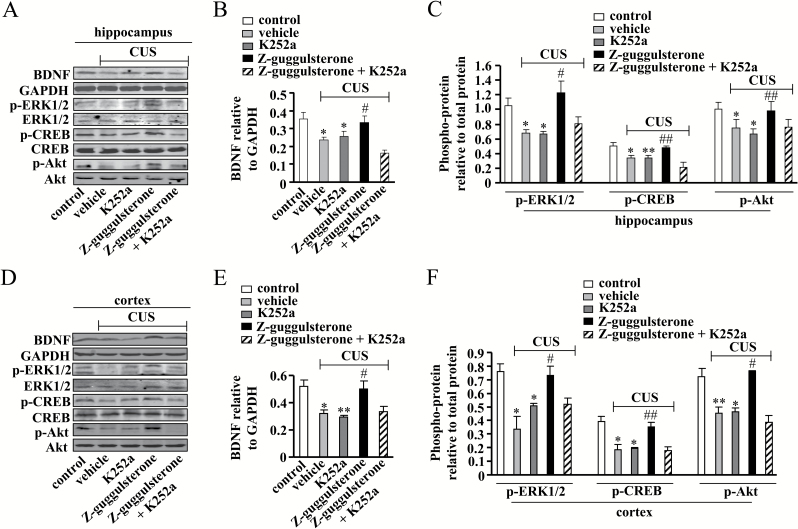
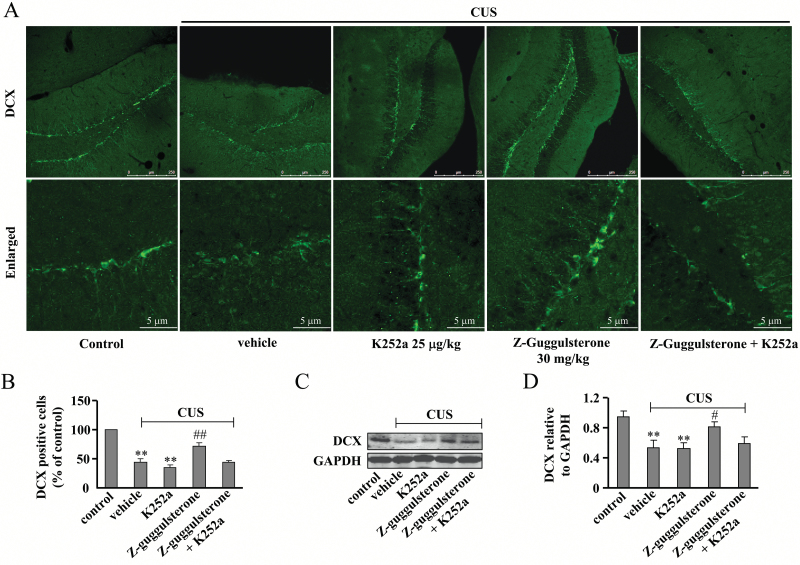
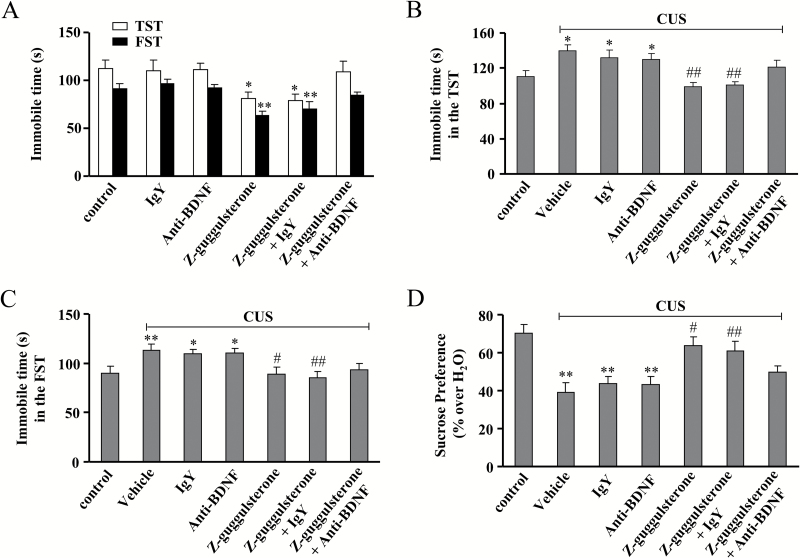
Results: Z-guggulsterone (10, 30 mg/kg) administration protected the mice against the chronic unpredictable stress-induced increases in the immobile time in the tail suspension test and forced swimming test and also reversed the reduction in sucrose intake in sucrose preference experiment. Z-guggulsterone (10, 30 mg/kg) administration prevented the reductions in brain-derived neurotrophic factor protein expression levels as well as the phosphorylation levels of cAMP response element binding protein, extracellular signal-regulated kinase 1/2, and protein kinase B in the hippocampus and cortex induced by chronic unpredictable stress. Z-guggulsterone (10, 30 mg/kg) treatment also improved hippocampal neurogenesis in chronic unpredictable stress-treated mice. Blockade of the brain-derived neurotrophic factor signal, but not the monoaminergic system, attenuated the antidepressant-like effects of Z-guggulsterone.
Conclusions: Z-guggulsterone exhibits antidepressant activity via activation of the brain-derived neurotrophic factor signaling pathway and upregulation of hippocampal neurogenesis.
Keywords: BDNF; CREB; CUS; Z-guggulsterone; major depression.
© The Author 2017. Published by Oxford University Press on behalf of CINP.
Figures
References
-
- Adams JD, Jr, Klaidman LK, Mishra L, Singh BB. (2002) Effects of guggul in a rat model of stroke. Altern Ther Health Med 8:20–21. - PubMed
-
- Barth M, Kriston L, Klostermann S, Barbui C, Cipriani A, Linde K. (2016) Efficacy of selective serotonin reuptake inhibitors and adverse events: meta-regression and mediation analysis of placebo-controlled trials. Br J Psychiatry 208:114−–119. - PubMed
-
- Berton O, Nestler EJ. (2006) New approaches to antidepressant drug discovery: beyond monoamines. Nat Rev Neurosci 7:137–151. - PubMed
-
- Bourin M, Fiocco AJ, Clenet F. (2001) How valuable are animal models in defining antidepressant activity? Hum Psychopharmacol 16:9–21. - PubMed
-
- Burke TF, Advani T, Adachi M, Monteggia LM, Hensler JG. (2013) Sensitivity of hippocampal 5-HT1A receptors to mild stress in BDNF-deficient mice. Int J Neuropsychopharmacol 16:631−–645. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
