

Gut symbiotic microbes imprint intestinal immune cells with the innate receptor SLAMF4 which contributes to gut immune protection against enteric pathogens
- PMID: 28341747
- PMCID: PMC5890651
- DOI: 10.1136/gutjnl-2016-313214
Gut symbiotic microbes imprint intestinal immune cells with the innate receptor SLAMF4 which contributes to gut immune protection against enteric pathogens
Abstract
Background: Interactions between host immune cells and gut microbiota are crucial for the integrity and function of the intestine. How these interactions regulate immune cell responses in the intestine remains a major gap in the field.
Aim: We have identified the signalling lymphocyte activation molecule family member 4 (SLAMF4) as an immunomodulator of the intestinal immunity. The aim is to determine how SLAMF4 is acquired in the gut and what its contribution to intestinal immunity is.
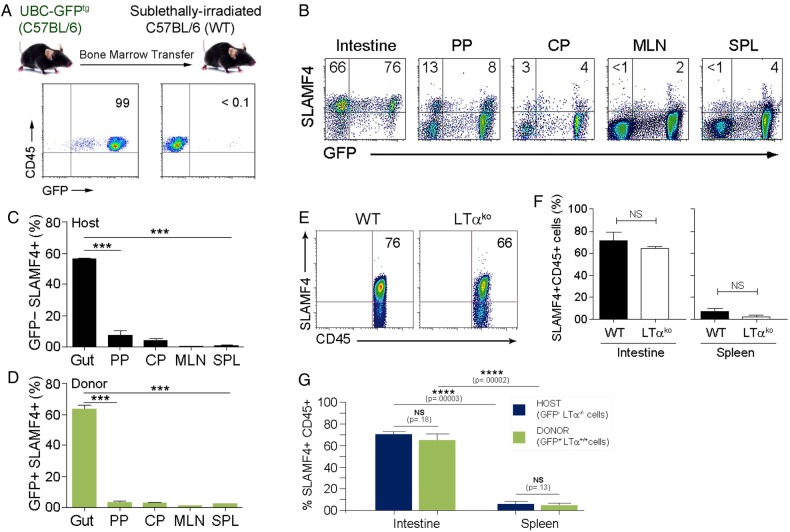
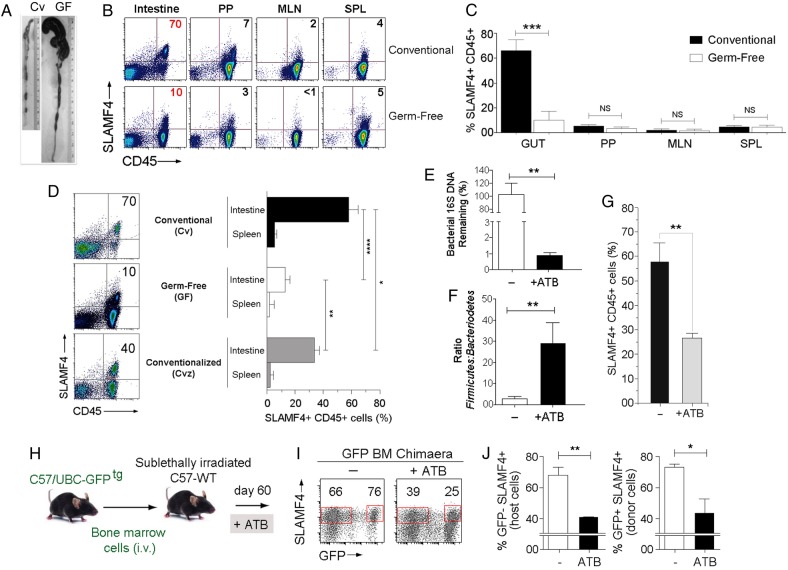
Methods: Expression of SLAMF4 was assessed in mice and humans. The mechanism of induction was studied using GFPtg bone marrow chimaera mice, lymphotoxin α and TNLG8A-deficient mice, as well as gnotobiotic mice. Role in immune protection was revealed using oral infection with Listeria monocytogenes and Cytobacter rodentium.
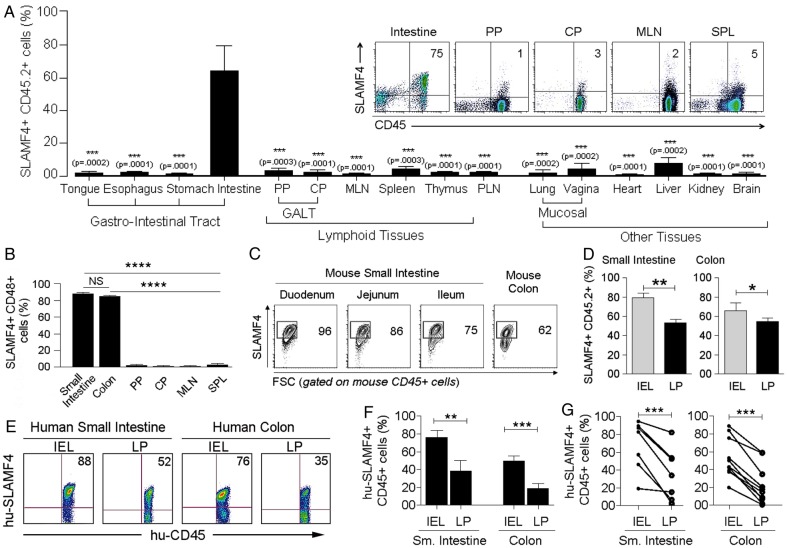
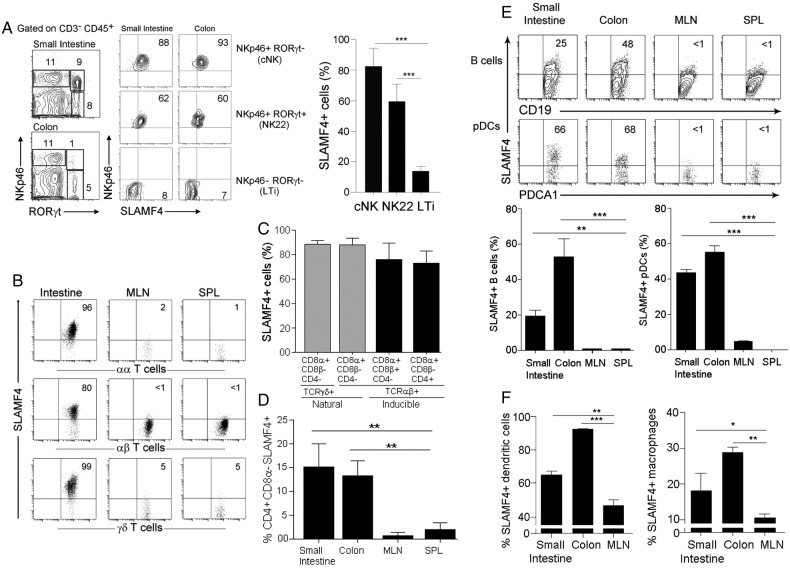
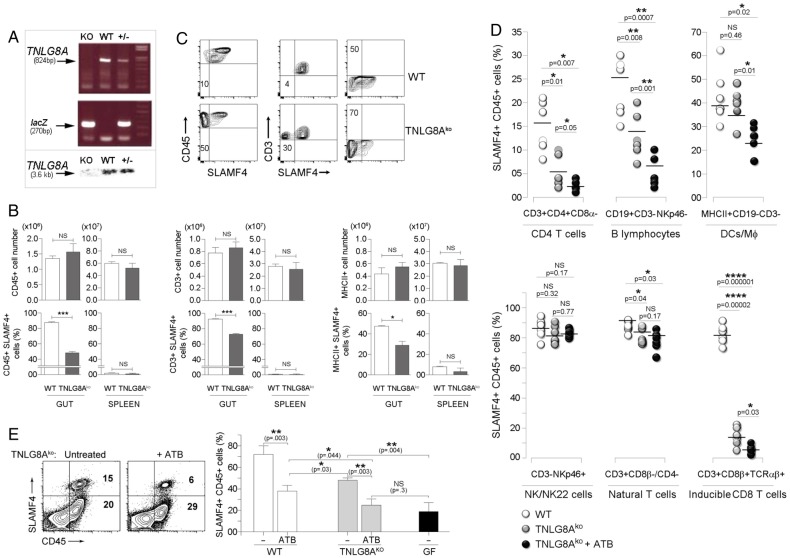
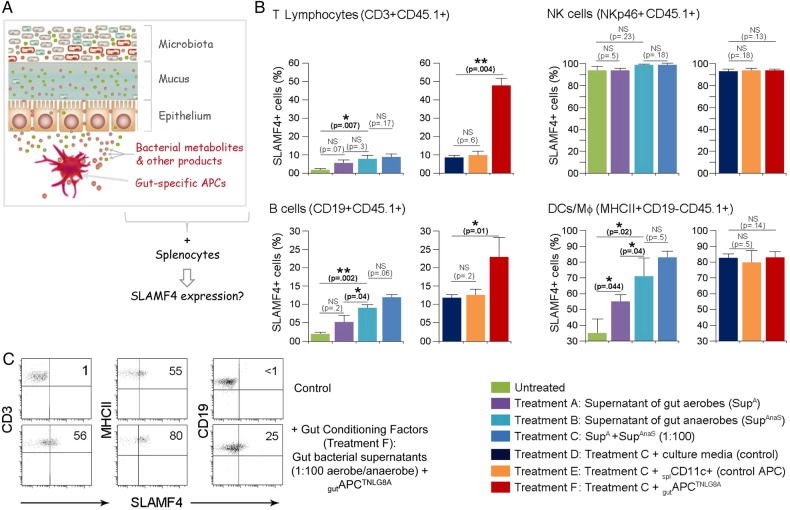
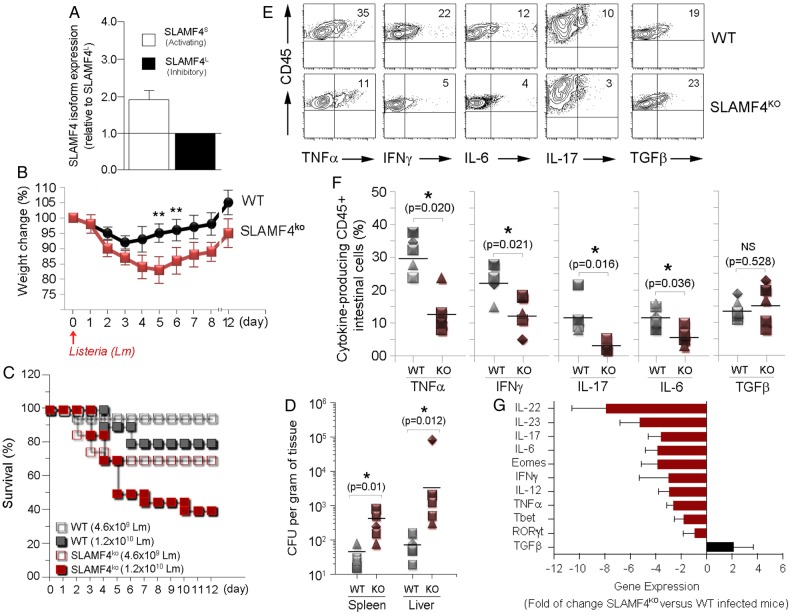
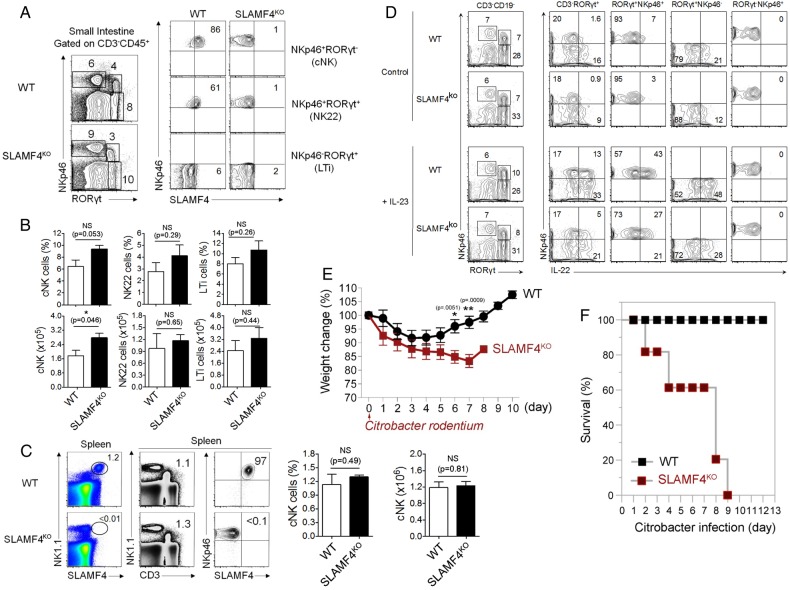
Results: SLAMF4 is a selective marker of intestinal immune cells of mice and humans. SLAMF4 induction occurs directly in the intestinal mucosa without the involvement of the gut-associated lymphoid tissue. Gut bacterial products, particularly those of gut anaerobes, and gut-resident antigen-presenting cell (APC) TNLG8A are key contributors of SLAMF4 induction in the intestine. Importantly, lack of SLAMF4 expression leads the increased susceptibility of mice to infection by oral pathogens culminating in their premature death.
Conclusions: SLAMF4 is a marker of intestinal immune cells which contributes to the protection against enteric pathogens and whose expression is dependent on the presence of the gut microbiota. This discovery provides a possible mechanism for answering the long-standing question of how the intertwining of the host and gut microbial biology regulates immune cell responses in the gut.
Keywords: ANTIBIOTICS; ENTERIC INFECTIONS; GASTROINTESTINAL IMMUNE RESPONSE; INTESTINAL BACTERIA; MUCOSAL IMMUNITY.
Published by the BMJ Publishing Group Limited. For permission to use (where not already granted under a licence) please go to http://www.bmj.com/company/products-services/rights-and-licensing/.
Conflict of interest statement
Competing interests: None declared.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous