

Uncovering specific changes in network wiring underlying the primate cerebrotype
- PMID: 28343248
- PMCID: PMC5585288
- DOI: 10.1007/s00429-017-1402-6
Uncovering specific changes in network wiring underlying the primate cerebrotype
Abstract
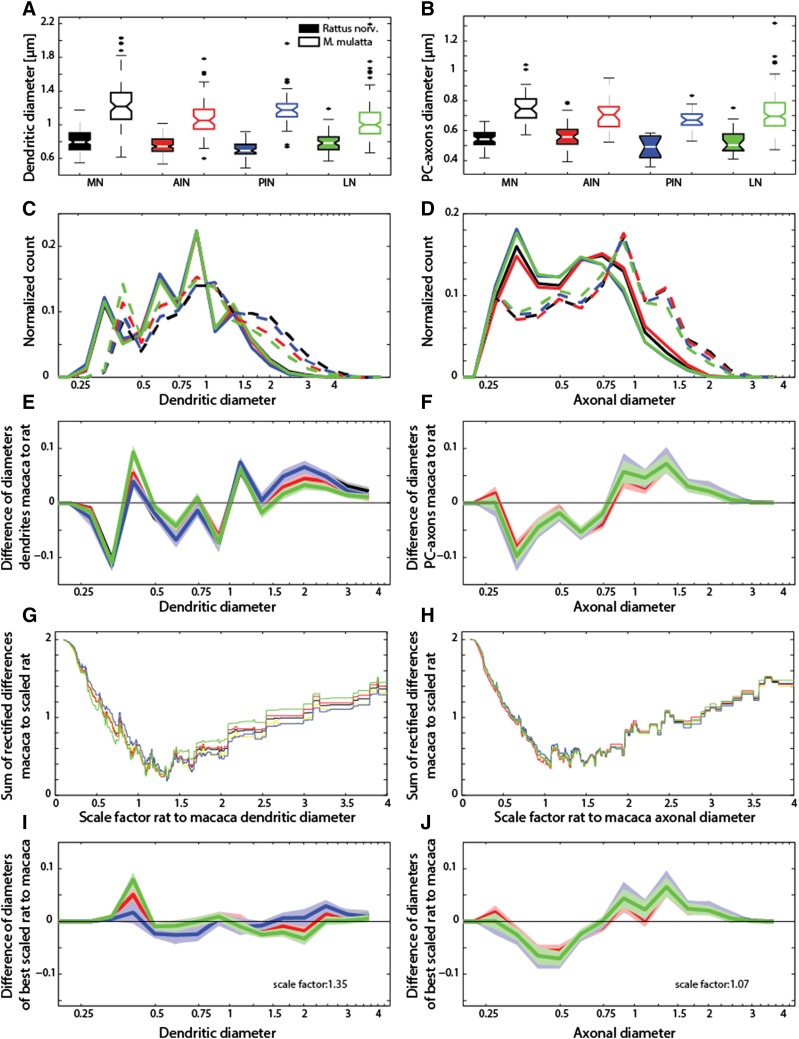
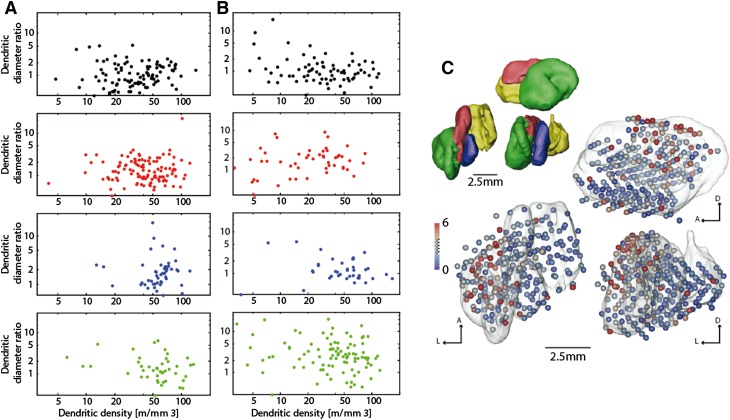
Regular scaling of brain networks during evolution has been proposed to be the major process leading to enlarged brains. Alternative views, however, suggest that deviations from regular scaling were crucial to the evolution of the primate brain and the emergence of different cerebrotypes. Here, we examined the scaling within the major link between the cerebellum and the cerebral cortex by studying the deep cerebellar nuclei (DCN). We compared the major axonal and dendritic wiring in the DCN of rodents and monkeys in search of regular scaling. We were able to confirm regular scaling within the density of neurons, the general dendritic length per neuron and the Purkinje cell axon length. However, we also observed specific modification of the scaling rules within the primates' largest and phylogenetically newest DCN, the dentate nucleus (LN/dentate). Our analysis shows a deviation from regular scaling in the predicted dendritic length per neuron in the LN/dentate. This reduction in the dendritic length is also associated with a smaller dendritic region-of-influence of these neurons. We also detected specific changes in the dendritic diameter distribution, supporting the theory that there is a shift in the neuronal population of the LN/dentate towards neurons that exhibit spatially restricted, clustered branching trees. The smaller dendritic fields would enable a larger number of network modules to be accommodated in the primate LN/dentate and would provide an explanation for the unique folded structure of the primate LN/dentate. Our results show that, in some brain regions, connectivity maximization (i.e., an increase of dendritic fields) is not the sole optimum and that increases in the number of network modules may be important for the emergence of a divergent primate cerebrotype.
Keywords: 3D reconstructions; Cerebellar nuclei; Comparative neuroanatomy; Dentate nucleus; Motor systems; Purkinje cells; Quantitative immunohistochemistry.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Figures

Similar articles
-
Quantitative organization of the excitatory synapses of the primate cerebellar nuclei: further evidence for a specialized architecture underlying the primate cerebellum.Brain Struct Funct. 2019 Jul;224(6):1987-1998. doi: 10.1007/s00429-019-01888-8. Epub 2019 May 17. Brain Struct Funct. 2019. PMID: 31101974 Free PMC article.
-
Systematic analysis of neuronal wiring of the rodent deep cerebellar nuclei reveals differences reflecting adaptations at the neuronal circuit and internuclear levels.J Comp Neurol. 2014 Aug 1;522(11):2481-97. doi: 10.1002/cne.23545. J Comp Neurol. 2014. PMID: 24477707
-
Axon collaterals of mossy fibers from the pontine nucleus in the cerebellar dentate nucleus.J Neurophysiol. 1992 Mar;67(3):547-60. doi: 10.1152/jn.1992.67.3.547. J Neurophysiol. 1992. PMID: 1578244
-
Not all brains are made the same: new views on brain scaling in evolution.Brain Behav Evol. 2011;78(1):22-36. doi: 10.1159/000327318. Epub 2011 Jun 17. Brain Behav Evol. 2011. PMID: 21691045 Review.
-
Cerebellar loops: a review of the nucleocortical pathway.Cerebellum. 2014 Jun;13(3):378-85. doi: 10.1007/s12311-013-0543-2. Cerebellum. 2014. PMID: 24362758 Free PMC article. Review.
Cited by
-
Learning-related contraction of gray matter in rodent sensorimotor cortex is associated with adaptive myelination.Elife. 2022 Nov 9;11:e77432. doi: 10.7554/eLife.77432. Elife. 2022. PMID: 36350292 Free PMC article.
-
Cerebrotypes in Cephalopods: Brain Diversity and Its Correlation With Species Habits, Life History, and Physiological Adaptations.Front Neuroanat. 2021 Feb 2;14:565109. doi: 10.3389/fnana.2020.565109. eCollection 2020. Front Neuroanat. 2021. PMID: 33603650 Free PMC article.
-
Quantitative organization of the excitatory synapses of the primate cerebellar nuclei: further evidence for a specialized architecture underlying the primate cerebellum.Brain Struct Funct. 2019 Jul;224(6):1987-1998. doi: 10.1007/s00429-019-01888-8. Epub 2019 May 17. Brain Struct Funct. 2019. PMID: 31101974 Free PMC article.
References
-
- Abeles M. Corticonics: neural circuits of the cerebral cortex. Cambridge: Cambridge University Press; 1991.
-
- Aizenman CD, Linden DJ. Regulation of the rebound depolarization and spontaneous firing patterns of deep nuclear neurons in slices of rat cerebellum. J Neurophysiol. 1999;82:1697–1709. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
