

Optimization of overexpression of a chaperone protein of steroid C25 dehydrogenase for biochemical and biophysical characterization
- PMID: 28343996
- PMCID: PMC5535313
- DOI: 10.1016/j.pep.2017.03.019
Optimization of overexpression of a chaperone protein of steroid C25 dehydrogenase for biochemical and biophysical characterization
Abstract
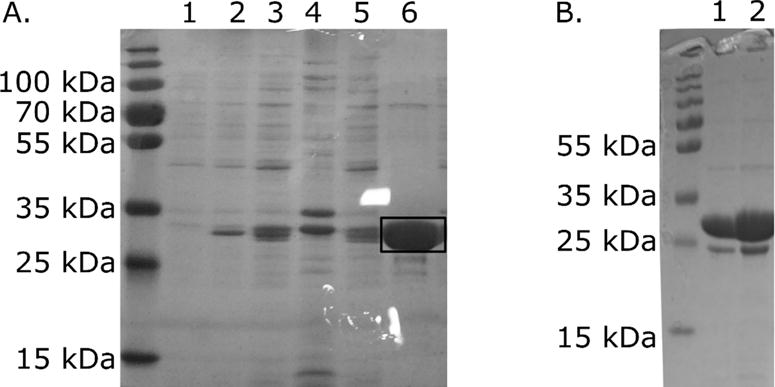
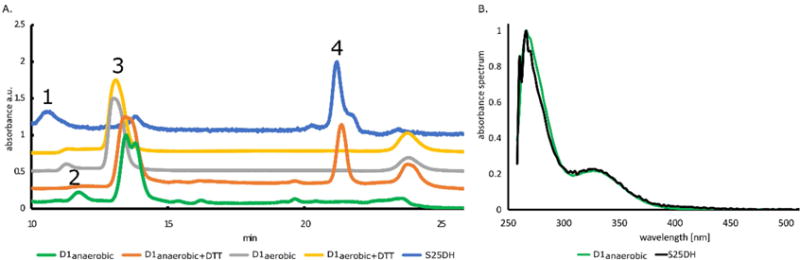
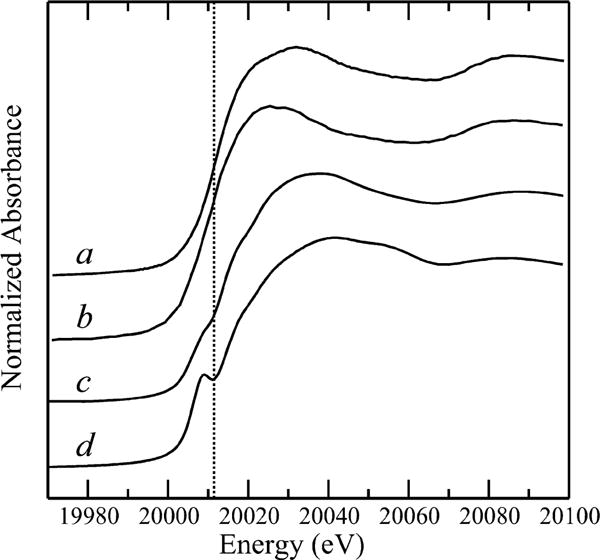
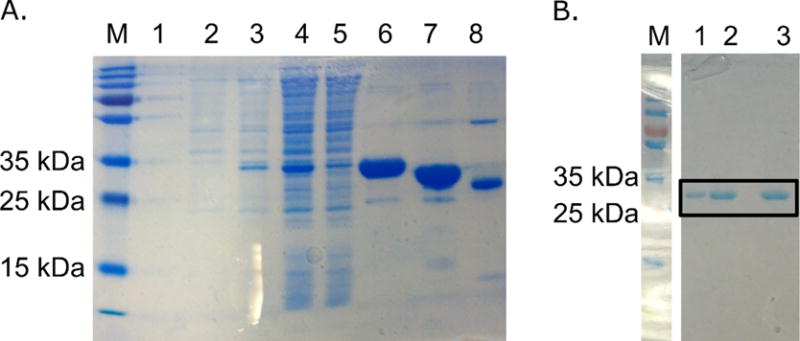
Molybdenum is an essential nutrient for metabolism in plant, bacteria, and animals. Molybdoenzymes are involved in nitrogen assimilation and oxidoreductive detoxification, and bioconversion reactions of environmental, industrial, and pharmaceutical interest. Molybdoenzymes contain a molybdenum cofactor (Moco), which is a pyranopterin heterocyclic compound that binds a molybdenum atom via a dithiolene group. Because Moco is a large and complex compound deeply buried within the protein, molybdoenzymes are accompanied by private chaperone proteins responsible for the cofactor's insertion into the enzyme and the enzyme's maturation. An efficient recombinant expression and purification of both Moco-free and Moco-containing molybdoenzymes and their chaperones is of paramount importance for fundamental and applied research related to molybdoenzymes. In this work, we focused on a D1 protein annotated as a chaperone of steroid C25 dehydrogenase (S25DH) from Sterolibacterium denitrificans Chol-1S. The D1 protein is presumably involved in the maturation of S25DH engaged in oxygen-independent oxidation of sterols. As this chaperone is thought to be a crucial element that ensures the insertion of Moco into the enzyme and consequently, proper folding of S25DH optimization of the chaperon's expression is the first step toward the development of recombinant expression and purification methods for S25DH. We have identified common E. coli strains and conditions for both expression and purification that allow us to selectively produce Moco-containing and Moco-free chaperones. We have also characterized the Moco-containing chaperone by EXAFS and HPLC analysis and identified conditions that stabilize both forms of the protein. The protocols presented here are efficient and result in protein quantities sufficient for biochemical studies.
Keywords: Chaperone protein; Molybdenum cofactor; Molybdoenzymes; Thermofluor shift assay.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures





Similar articles
-
The biosynthesis of the molybdenum cofactors in Escherichia coli.Environ Microbiol. 2020 Jun;22(6):2007-2026. doi: 10.1111/1462-2920.15003. Epub 2020 Apr 6. Environ Microbiol. 2020. PMID: 32239579 Review.
-
Molybdenum enzymes, their maturation and molybdenum cofactor biosynthesis in Escherichia coli.Biochim Biophys Acta. 2013 Aug-Sep;1827(8-9):1086-101. doi: 10.1016/j.bbabio.2012.11.007. Epub 2012 Nov 29. Biochim Biophys Acta. 2013. PMID: 23201473 Review.
-
The role of FeS clusters for molybdenum cofactor biosynthesis and molybdoenzymes in bacteria.Biochim Biophys Acta. 2015 Jun;1853(6):1335-49. doi: 10.1016/j.bbamcr.2014.09.021. Epub 2014 Sep 28. Biochim Biophys Acta. 2015. PMID: 25268953 Free PMC article. Review.
-
The biosynthesis of the molybdenum cofactors.J Biol Inorg Chem. 2015 Mar;20(2):337-47. doi: 10.1007/s00775-014-1173-y. Epub 2014 Jul 1. J Biol Inorg Chem. 2015. PMID: 24980677
-
Transfer of the molybdenum cofactor synthesized by Rhodobacter capsulatus MoeA to XdhC and MobA.J Biol Chem. 2007 Sep 28;282(39):28493-28500. doi: 10.1074/jbc.M704020200. Epub 2007 Aug 7. J Biol Chem. 2007. PMID: 17686778
Cited by
-
Characterizing metal-binding sites in proteins with X-ray crystallography.Nat Protoc. 2018 May;13(5):1062-1090. doi: 10.1038/nprot.2018.018. Epub 2018 Apr 19. Nat Protoc. 2018. PMID: 29674755 Free PMC article.
-
Bacterial steroid hydroxylases: enzyme classes, their functions and comparison of their catalytic mechanisms.Appl Microbiol Biotechnol. 2018 Oct;102(19):8153-8171. doi: 10.1007/s00253-018-9239-3. Epub 2018 Jul 21. Appl Microbiol Biotechnol. 2018. PMID: 30032434 Free PMC article. Review.
-
History of Maturation of Prokaryotic Molybdoenzymes-A Personal View.Molecules. 2023 Oct 20;28(20):7195. doi: 10.3390/molecules28207195. Molecules. 2023. PMID: 37894674 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
