

Effective Depletion of Pre-existing Anti-AAV Antibodies Requires Broad Immune Targeting
- PMID: 28345001
- PMCID: PMC5363314
- DOI: 10.1016/j.omtm.2017.01.003
Effective Depletion of Pre-existing Anti-AAV Antibodies Requires Broad Immune Targeting
Abstract
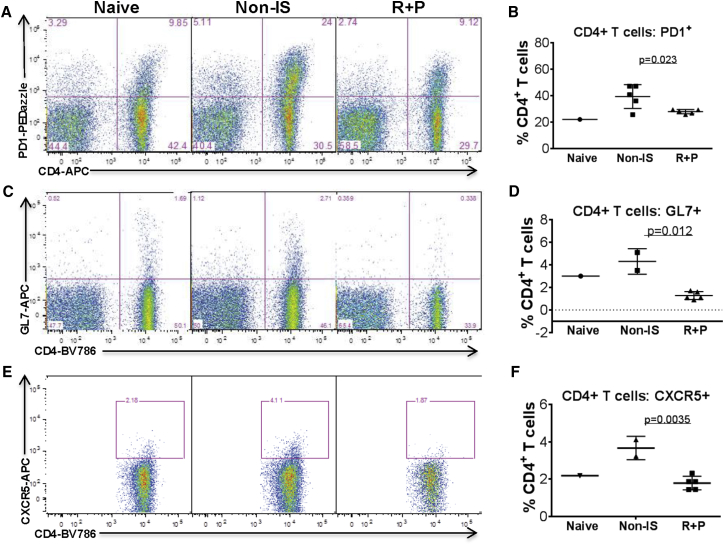
Pre-existing antibodies (Abs) to AAV pose a critical challenge for the translation of gene therapies. No effective approach is available to overcome pre-existing Abs. Given the complexity of Ab production, overcoming pre-existing Abs will require broad immune targeting. We generated a mouse model of pre-existing AAV9 Abs to test multiple immunosuppressants, including bortezomib, rapamycin, and prednisolone, individually or in combination. We identified an effective approach combining rapamycin and prednisolone, reducing serum AAV9 Abs by 70%-80% at 4 weeks and 85%-93% at 8 weeks of treatment. The rapamycin plus prednisolone treatment resulted in significant decreases in the frequency of B cells, plasma cells, and IgG-secreting and AAV9-specific Ab-producing plasma cells in bone marrow. The rapamycin plus prednisolone treatment also significantly reduced frequencies of IgD-IgG+ class-switched/FAS+CL7+ germinal center B cells, and of activated CD4+ T cells expressing PD1 and GL7, in spleen. These data suggest that rapamycin plus prednisolone has selective inhibitory effects on both T helper type 2 support of B cell activation in spleen and on bone marrow plasma cell survival, leading to effective AAV9 Abs depletion. This promising immunomodulation approach is highly translatable, and it poses minimal risk in the context of therapeutic benefits promised by gene therapy for severe monogenetic diseases, with a single or possibly a few treatments over a lifetime.
Keywords: AAV; gene therapy; pre-existing antibody.
Figures





References
-
- Manno C.S., Pierce G.F., Arruda V.R., Glader B., Ragni M., Rasko J.J., Ozelo M.C., Hoots K., Blatt P., Konkle B. Successful transduction of liver in hemophilia by AAV-Factor IX and limitations imposed by the host immune response. Nat. Med. 2006;12:342–347. - PubMed
-
- Nathwani A.C., Rosales C., McIntosh J., Rastegarlari G., Nathwani D., Raj D., Nawathe S., Waddington S.N., Bronson R., Jackson S. Long-term safety and efficacy following systemic administration of a self-complementary AAV vector encoding human FIX pseudotyped with serotype 5 and 8 capsid proteins. Mol. Ther. 2011;19:876–885. - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
