

Insights into electrosensory organ development, physiology and evolution from a lateral line-enriched transcriptome
- PMID: 28346141
- PMCID: PMC5429088
- DOI: 10.7554/eLife.24197
Insights into electrosensory organ development, physiology and evolution from a lateral line-enriched transcriptome
Abstract
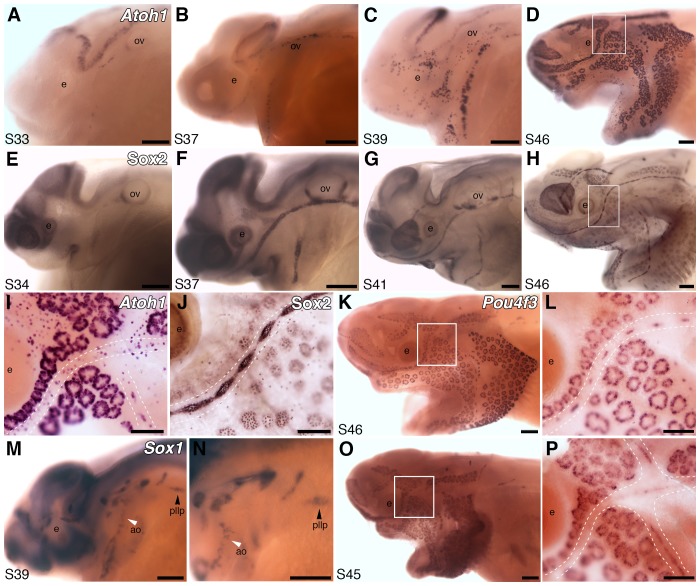
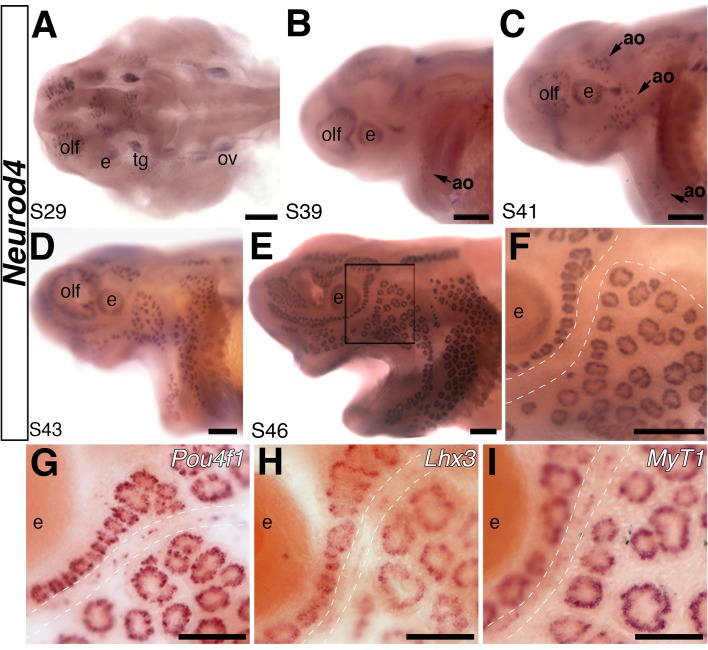
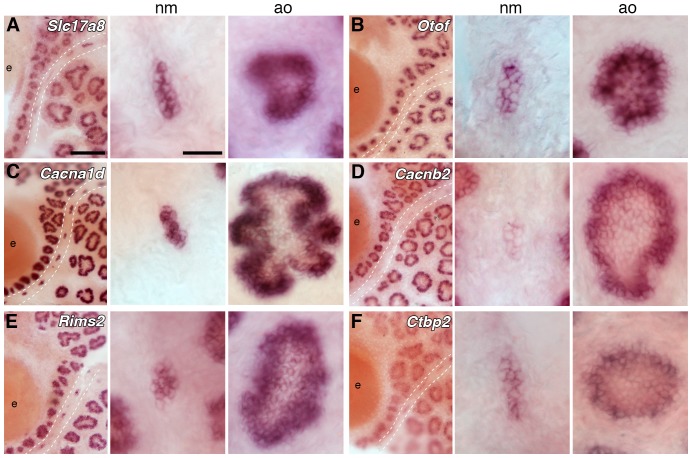
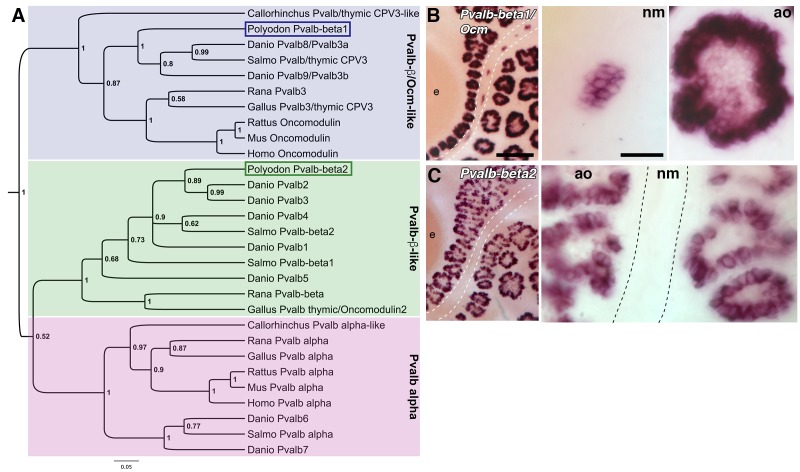
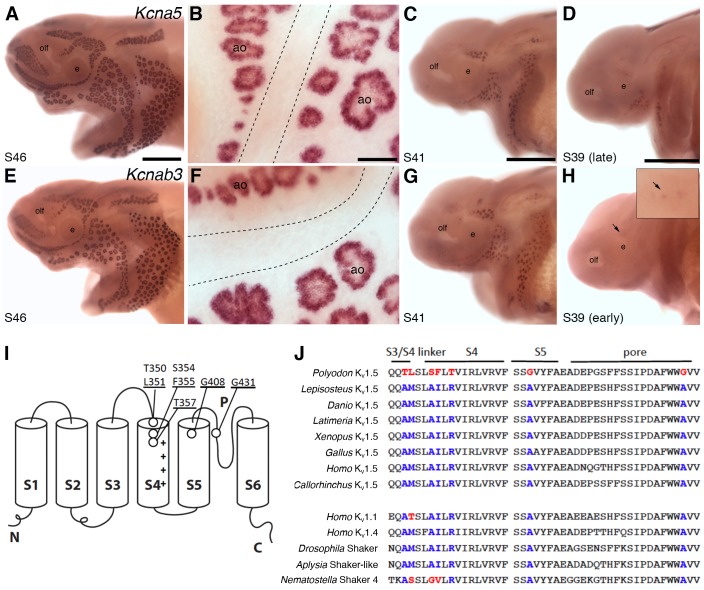
The anamniote lateral line system, comprising mechanosensory neuromasts and electrosensory ampullary organs, is a useful model for investigating the developmental and evolutionary diversification of different organs and cell types. Zebrafish neuromast development is increasingly well understood, but neither zebrafish nor Xenopus is electroreceptive and our molecular understanding of ampullary organ development is rudimentary. We have used RNA-seq to generate a lateral line-enriched gene-set from late-larval paddlefish (Polyodon spathula). Validation of a subset reveals expression in developing ampullary organs of transcription factor genes critical for hair cell development, and genes essential for glutamate release at hair cell ribbon synapses, suggesting close developmental, physiological and evolutionary links between non-teleost electroreceptors and hair cells. We identify an ampullary organ-specific proneural transcription factor, and candidates for the voltage-sensing L-type Cav channel and rectifying Kv channel predicted from skate (cartilaginous fish) ampullary organ electrophysiology. Overall, our results illuminate ampullary organ development, physiology and evolution.
Keywords: Atoh1; Cav1.3; Cavβ2; Kv1.5; Kvβ3; Neurod4; Polyodon spathula (Mississippi paddlefish); Pou4f3; Vglut3; ampullary organs; beta-parvalbumins; developmental biology; electroreceptors; hair cells; hh; neuromasts; neuroscience; oncomodulin; otoferlin; stem cells; synaptic ribbons; voltage-gated ion channels.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
References
-
- Alves-Gomes JA. The evolution of electroreception and bioelectrogenesis in teleost fish: a phylogenetic perspective. Journal of Fish Biology. 2001;58:1489–1511. doi: 10.1111/j.1095-8649.2001.tb02307.x. - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
