

A genetic switch controls the production of flagella and toxins in Clostridium difficile
- PMID: 28346491
- PMCID: PMC5386303
- DOI: 10.1371/journal.pgen.1006701
A genetic switch controls the production of flagella and toxins in Clostridium difficile
Abstract
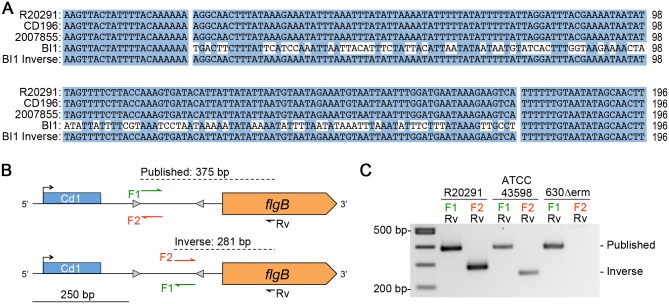
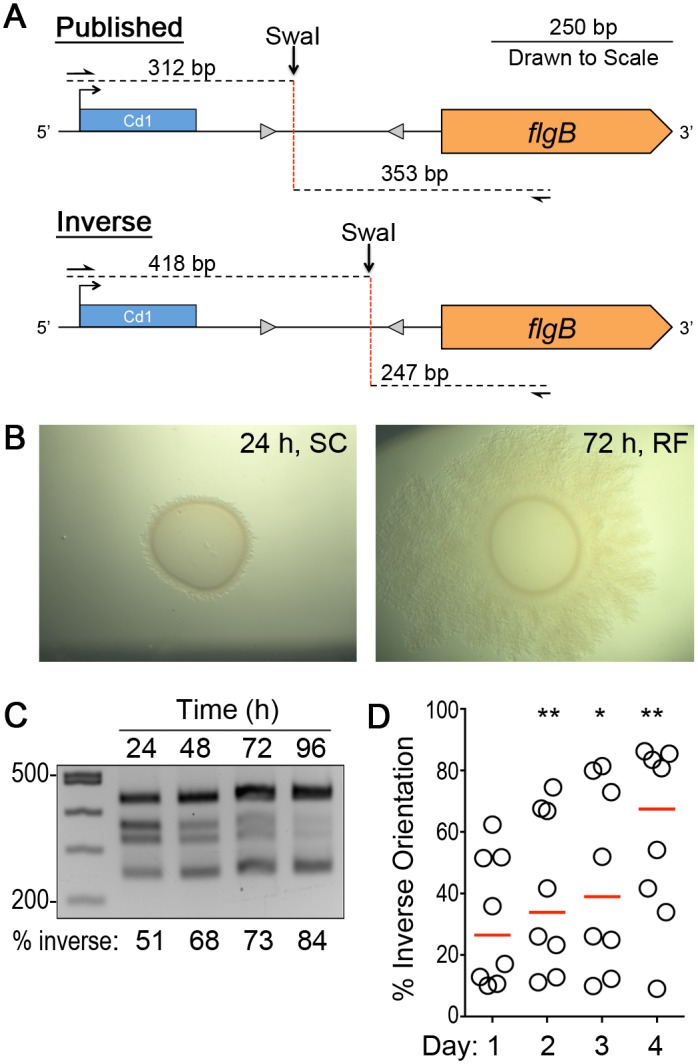
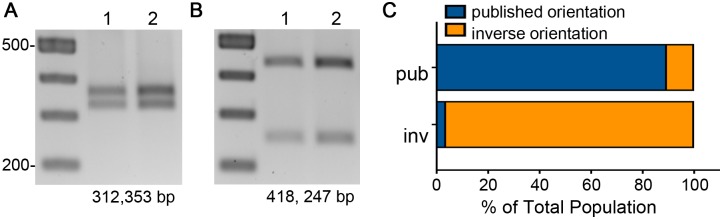
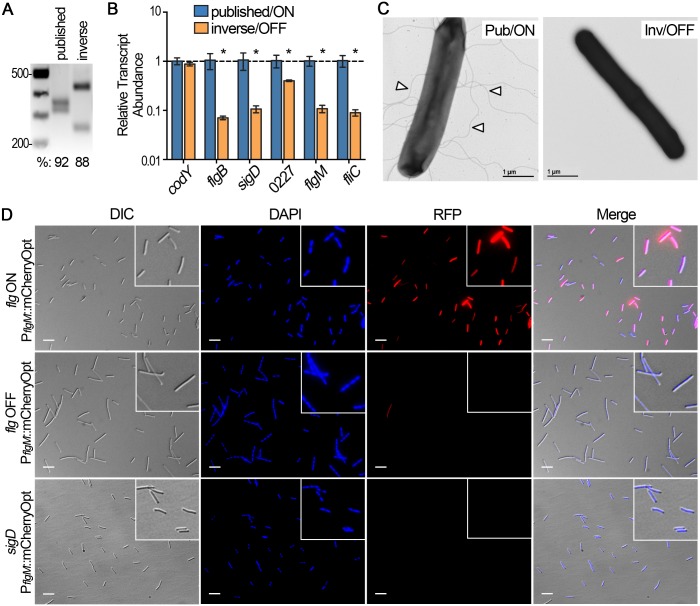
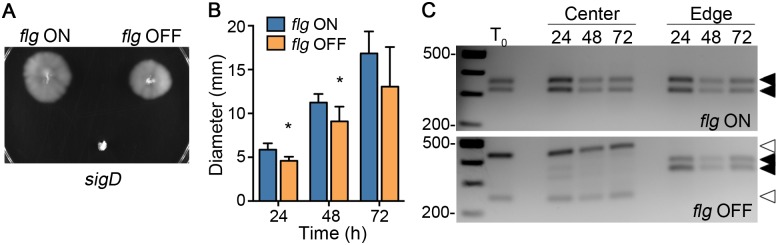
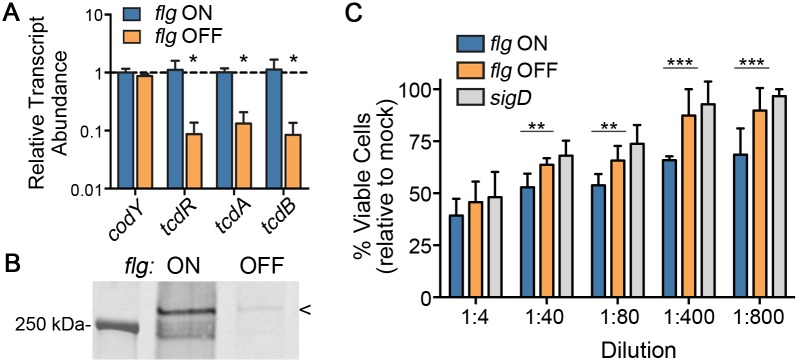
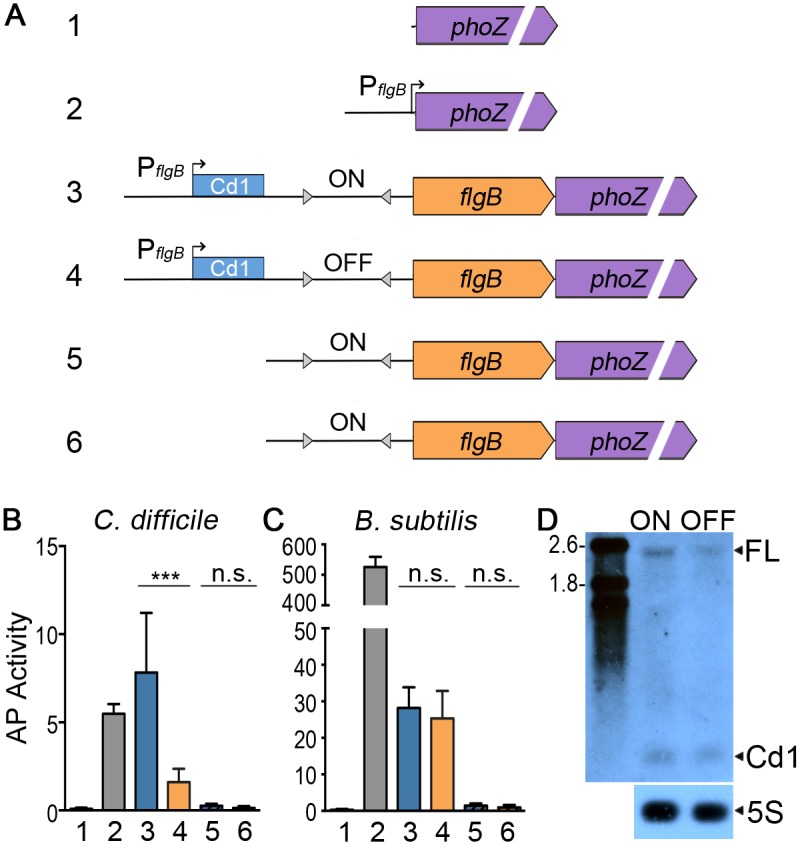
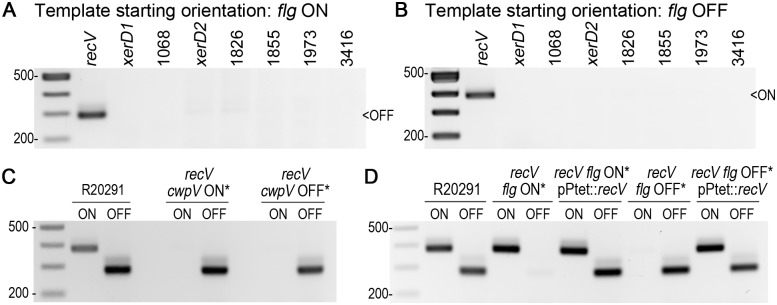
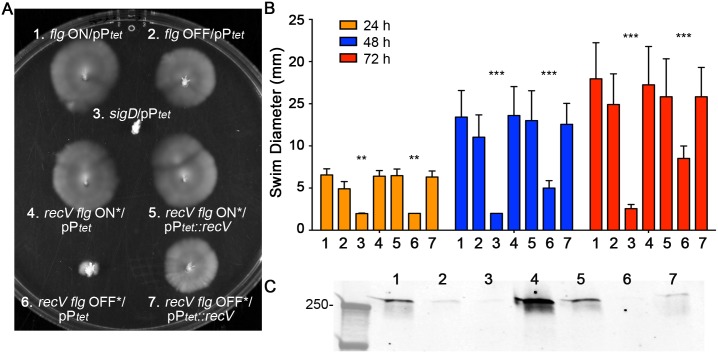
In the human intestinal pathogen Clostridium difficile, flagella promote adherence to intestinal epithelial cells. Flagellar gene expression also indirectly impacts production of the glucosylating toxins, which are essential to diarrheal disease development. Thus, factors that regulate the expression of the flgB operon will likely impact toxin production in addition to flagellar motility. Here, we report the identification a "flagellar switch" that controls the phase variable production of flagella and glucosylating toxins. The flagellar switch, located upstream of the flgB operon containing the early stage flagellar genes, is a 154 bp invertible sequence flanked by 21 bp inverted repeats. Bacteria with the sequence in one orientation expressed flagellum and toxin genes, produced flagella, and secreted the toxins ("flg phase ON"). Bacteria with the sequence in the inverse orientation were attenuated for flagellar and toxin gene expression, were aflagellate, and showed decreased toxin secretion ("flg phase OFF"). The orientation of the flagellar switch is reversible during growth in vitro. We provide evidence that gene regulation via the flagellar switch occurs post-transcription initiation and requires a C. difficile-specific regulatory factor to destabilize or degrade the early flagellar gene mRNA when the flagellar switch is in the OFF orientation. Lastly, through mutagenesis and characterization of flagellar phase locked isolates, we determined that the tyrosine recombinase RecV, which catalyzes inversion at the cwpV switch, is also responsible for inversion at the flagellar switch in both directions. Phase variable flagellar motility and toxin production suggests that these important virulence factors have both advantageous and detrimental effects during the course of infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
