

Low Antigen Dose in Adjuvant-Based Vaccination Selectively Induces CD4 T Cells with Enhanced Functional Avidity and Protective Efficacy
- PMID: 28348274
- PMCID: PMC5392729
- DOI: 10.4049/jimmunol.1600965
Low Antigen Dose in Adjuvant-Based Vaccination Selectively Induces CD4 T Cells with Enhanced Functional Avidity and Protective Efficacy
Abstract
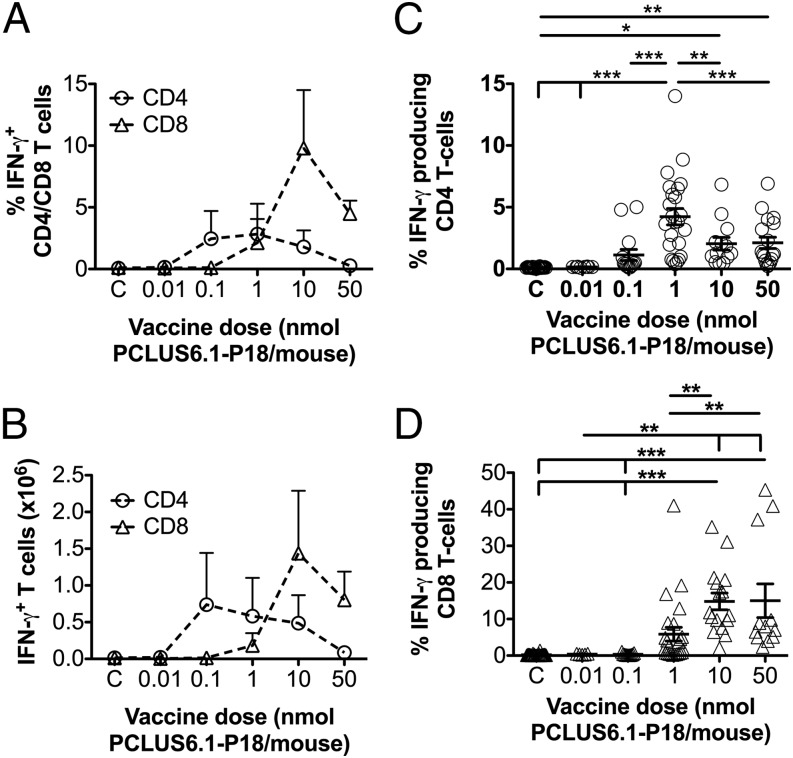
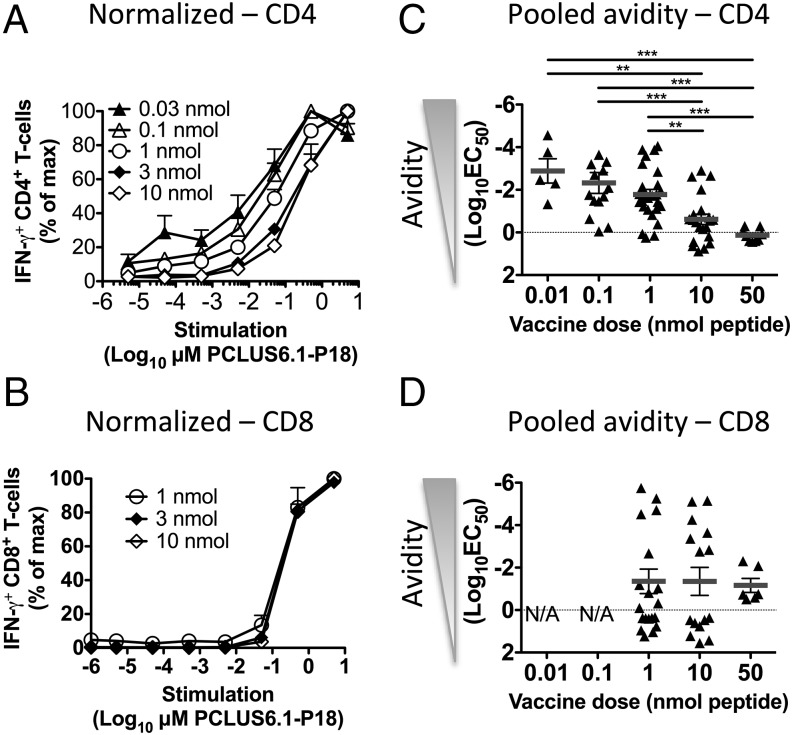
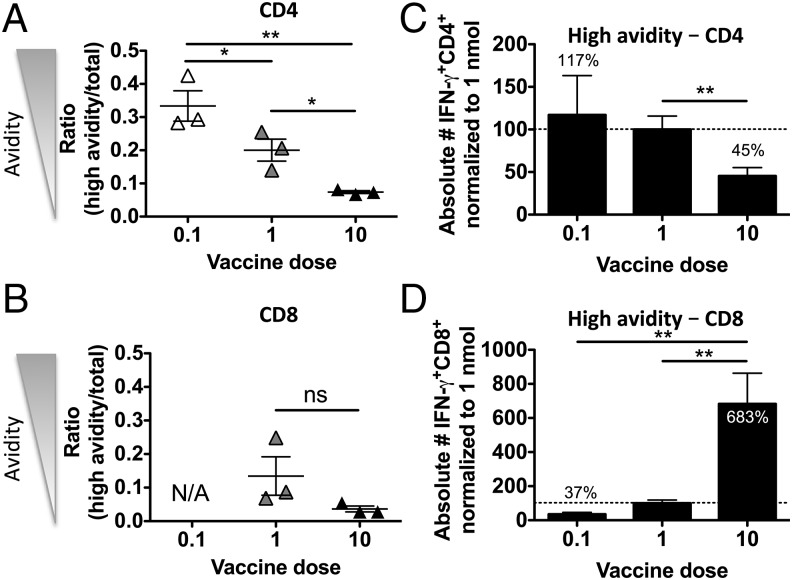
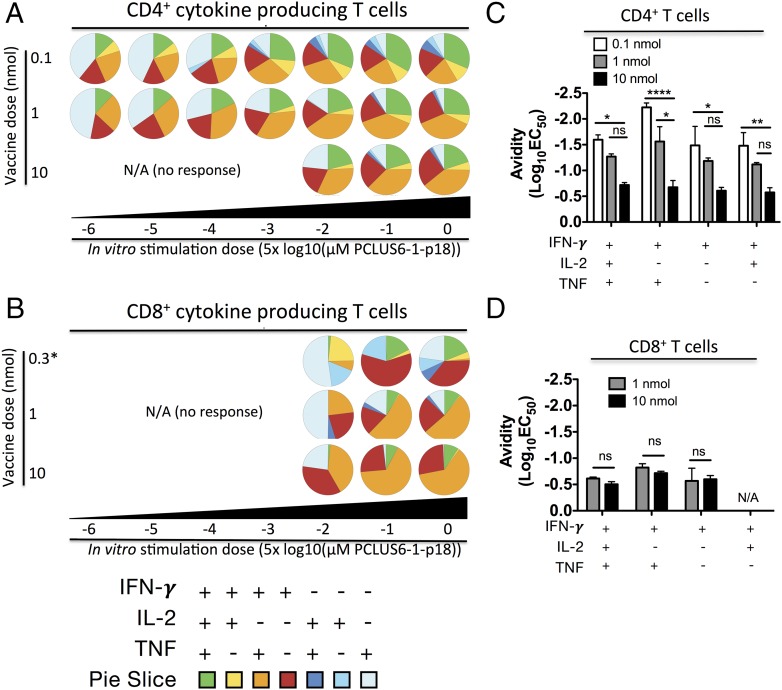
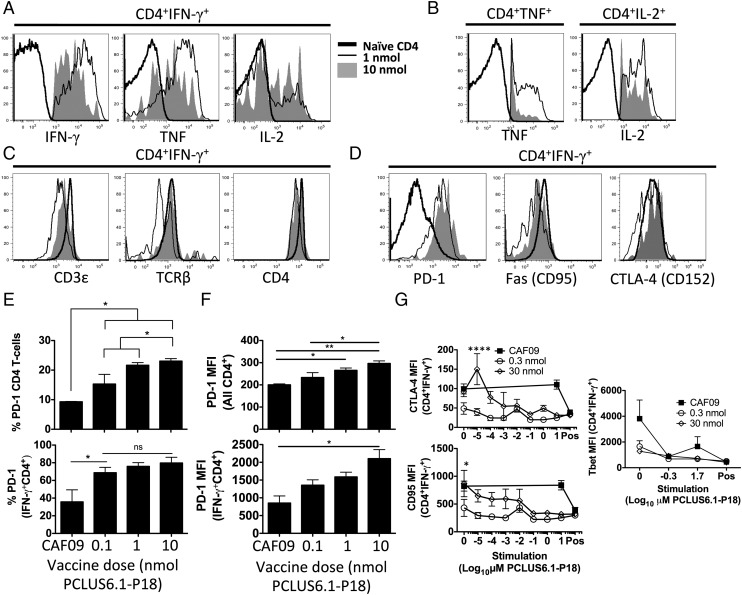
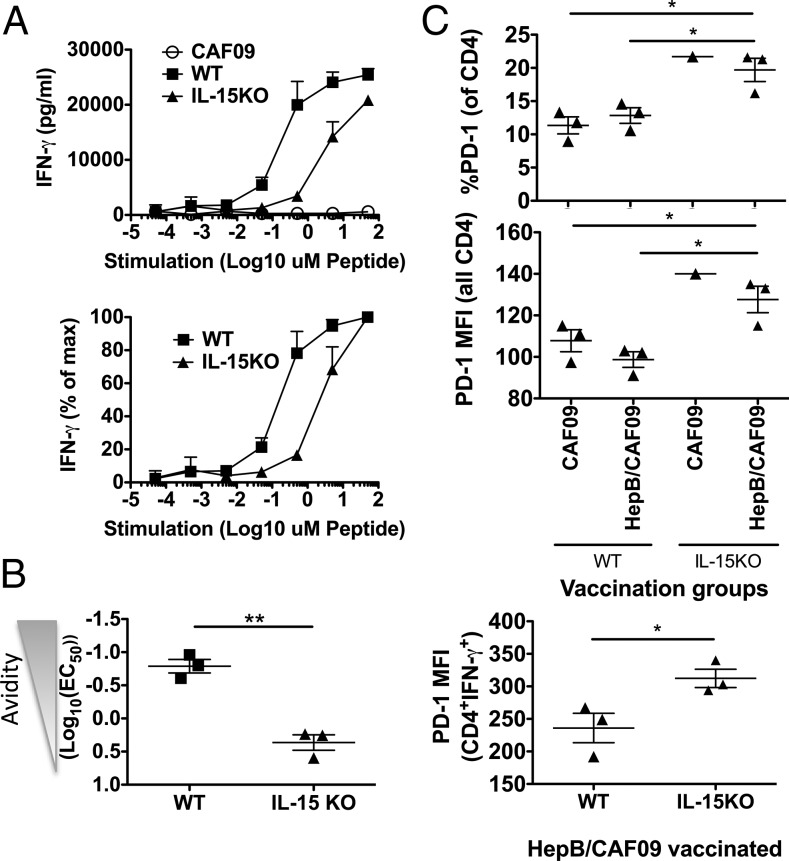
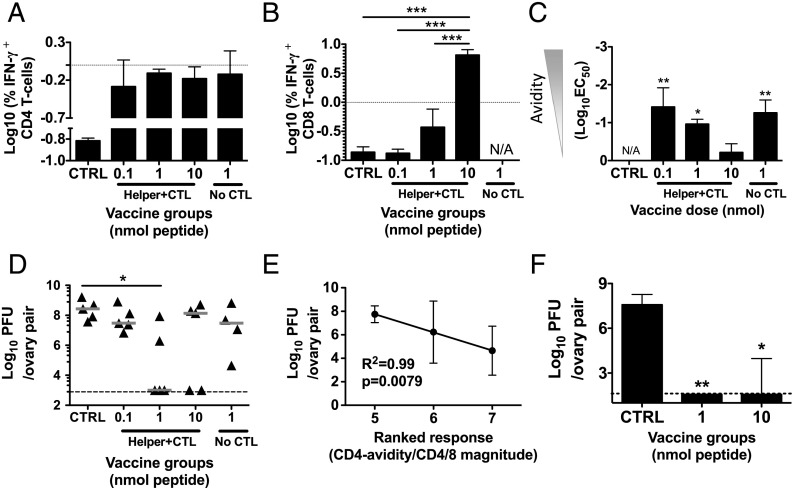
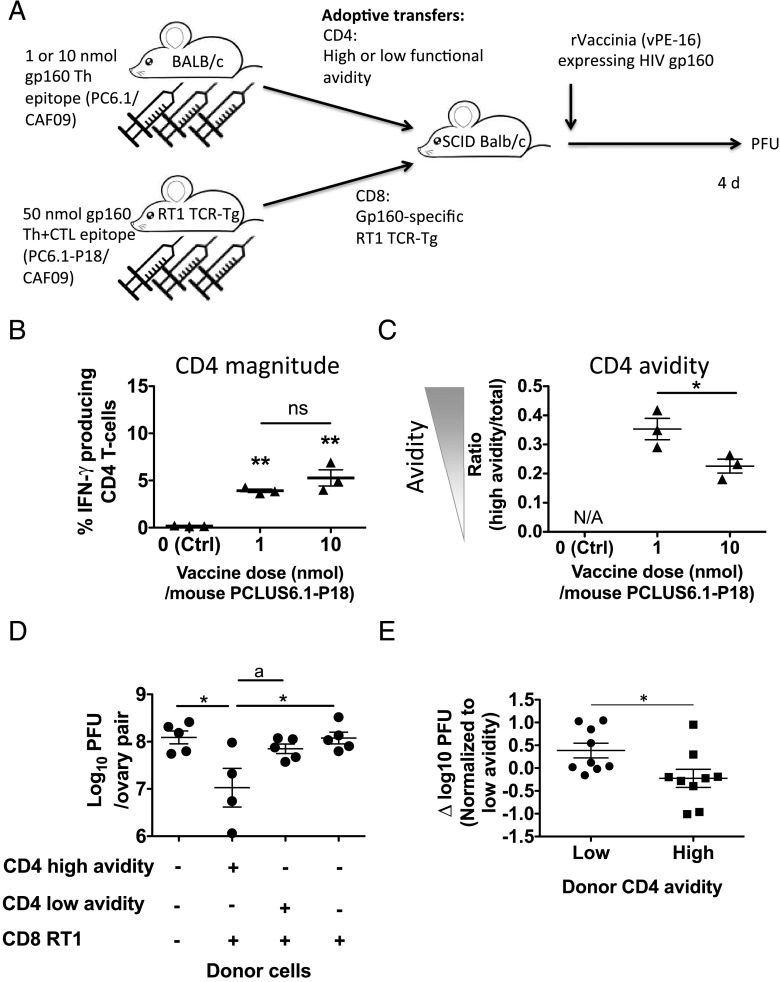
T cells with high functional avidity can sense and respond to low levels of cognate Ag, a characteristic that is associated with more potent responses against tumors and many infections, including HIV. Although an important determinant of T cell efficacy, it has proven difficult to selectively induce T cells of high functional avidity through vaccination. Attempts to induce high-avidity T cells by low-dose in vivo vaccination failed because this strategy simply gave no response. Instead, selective induction of high-avidity T cells has required in vitro culturing of specific T cells with low Ag concentrations. In this study, we combined low vaccine Ag doses with a novel potent cationic liposomal adjuvant, cationic adjuvant formulation 09, consisting of dimethyldioctadecylammonium liposomes incorporating two immunomodulators (monomycolyl glycerol analog and polyinosinic-polycytidylic acid) that efficiently induces CD4 Th cells, as well as cross-primes CD8 CTL responses. We show that vaccination with low Ag dose selectively primes CD4 T cells of higher functional avidity, whereas CD8 T cell functional avidity was unrelated to vaccine dose in mice. Importantly, CD4 T cells of higher functional avidity induced by low-dose vaccinations showed higher cytokine release per cell and lower inhibitory receptor expression (PD-1, CTLA-4, and the apoptosis-inducing Fas death receptor) compared with their lower-avidity CD4 counterparts. Notably, increased functional CD4 T cell avidity improved antiviral efficacy of CD8 T cells. These data suggest that potent adjuvants, such as cationic adjuvant formulation 09, render low-dose vaccination a feasible and promising approach for generating high-avidity T cells through vaccination.
Copyright © 2017 by The American Association of Immunologists, Inc.
Figures
References
-
- Seder R. A., Darrah P. A., Roederer M. 2008. T-cell quality in memory and protection: implications for vaccine design. Nat. Rev. Immunol. 8: 247–258. - PubMed
-
- Bullock T. N., Mullins D. W., Engelhard V. H. 2003. Antigen density presented by dendritic cells in vivo differentially affects the number and avidity of primary, memory, and recall CD8+ T cells. J. Immunol. 170: 1822–1829. - PubMed
-
- Derby M., Alexander-Miller M., Tse R., Berzofsky J. 2001. High-avidity CTL exploit two complementary mechanisms to provide better protection against viral infection than low-avidity CTL. J. Immunol. 166: 1690–1697. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
