

Cooperation and competition shape ecological resistance during periodic spatial disturbance of engineered bacteria
- PMID: 28348396
- PMCID: PMC5428654
- DOI: 10.1038/s41598-017-00588-9
Cooperation and competition shape ecological resistance during periodic spatial disturbance of engineered bacteria
Abstract
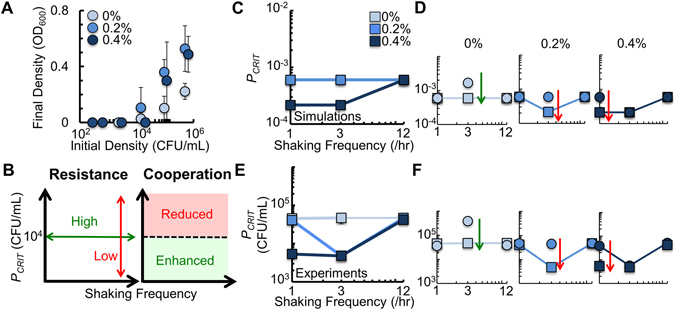
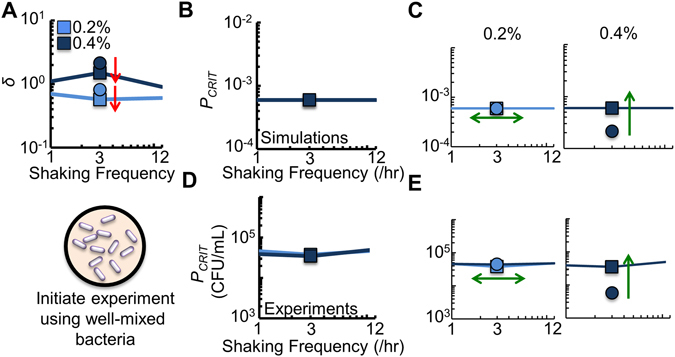
Cooperation is fundamental to the survival of many bacterial species. Previous studies have shown that spatial structure can both promote and suppress cooperation. Most environments where bacteria are found are periodically disturbed, which can affect the spatial structure of the population. Despite the important role that spatial disturbances play in maintaining ecological relationships, it remains unclear as to how periodic spatial disturbances affect bacteria dependent on cooperation for survival. Here, we use bacteria engineered with a strong Allee effect to investigate how the frequency of periodic spatial disturbances affects cooperation. We show that at intermediate frequencies of spatial disturbance, the ability of the bacterial population to cooperate is perturbed. A mathematical model demonstrates that periodic spatial disturbance leads to a tradeoff between accessing an autoinducer and accessing nutrients, which determines the ability of the bacteria to cooperate. Based on this relationship, we alter the ability of the bacteria to access an autoinducer. We show that increased access to an autoinducer can enhance cooperation, but can also reduce ecological resistance, defined as the ability of a population to resist changes due to disturbance. Our results may have implications in maintaining stability of microbial communities and in the treatment of infectious diseases.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
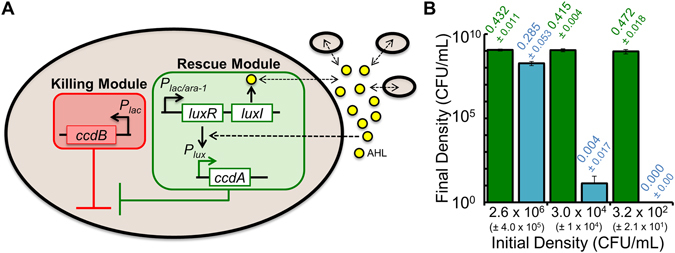
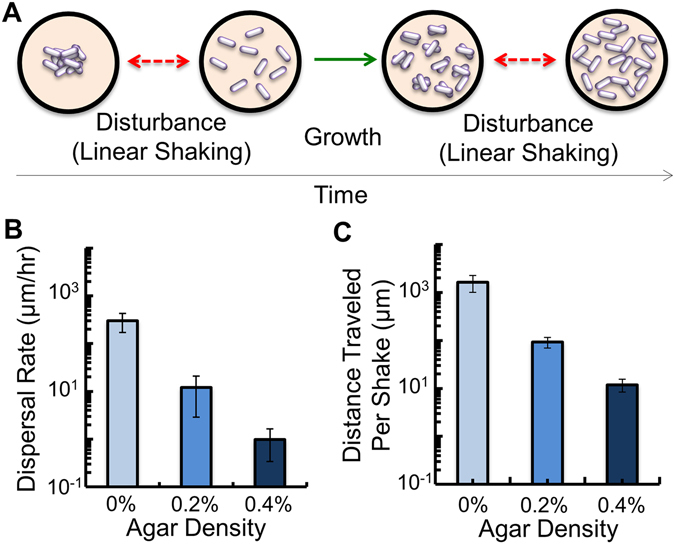
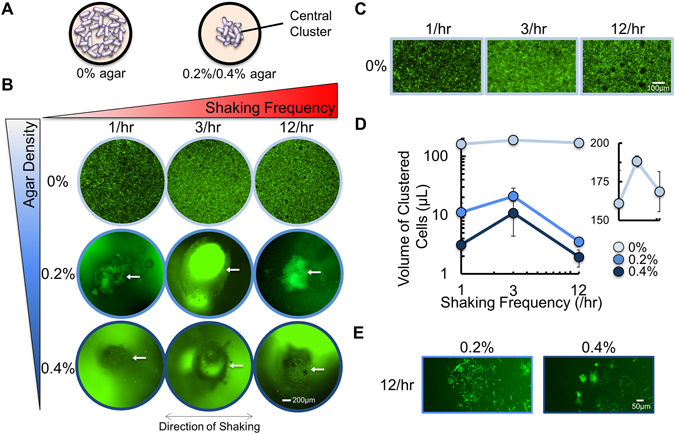
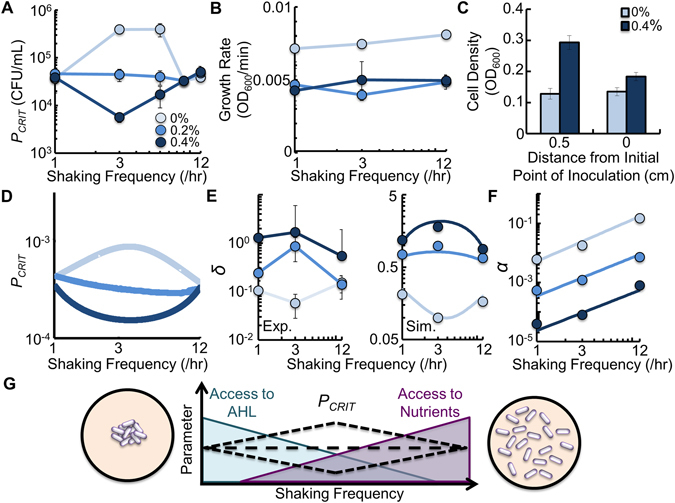
References
-
- Redfield, R. J. Is quorum sensing a side effect of diffusion sensing? Trends in Microbiology10, 365–370, doi:http://dx.doi.org/10.1016/S0966-842X(02)02400-9 (2002). - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
