

Organization of prefrontal network activity by respiration-related oscillations
- PMID: 28349959
- PMCID: PMC5368652
- DOI: 10.1038/srep45508
Organization of prefrontal network activity by respiration-related oscillations
Abstract
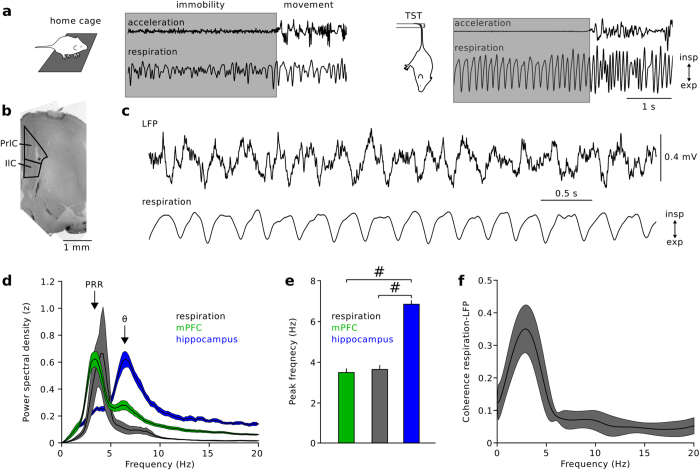
The medial prefrontal cortex (mPFC) integrates information from cortical and sub-cortical areas and contributes to the planning and initiation of behaviour. A potential mechanism for signal integration in the mPFC lies in the synchronization of neuronal discharges by theta (6-12 Hz) activity patterns. Here we show, using in vivo local field potential (LFP) and single-unit recordings from awake mice, that prominent oscillations in the sub-theta frequency band (1-5 Hz) emerge during awake immobility in the mPFC. These oscillation patterns are distinct from but phase-locked to hippocampal theta activity and occur synchronized with nasal respiration (hence termed prefrontal respiration rhythm [PRR]). PRR activity modulates the amplitude of prefrontal gamma rhythms with greater efficacy than theta oscillations. Furthermore, single-unit discharges of putative pyramidal cells and GABAergic interneurons are entrained by prefrontal PRR and nasal respiration. Our data thus suggest that PRR activity contributes to information processing in the prefrontal neuronal network.
Conflict of interest statement
The authors declare no competing financial interests.
Figures




References
-
- Lee M. G., Chrobak J. J., Sik A., Wiley R. G. & Buzsáki G. Hippocampal theta activity following selective lesion of the septal cholinergic system. Neuroscience 62, 1033–1047 (1994). - PubMed
-
- Wang X.-J. Pacemaker neurons for the theta rhythm and their synchronization in the septohippocampal reciprocal loop. J. Neurophysiol. 87, 889–900 (2002). - PubMed
-
- Dégenètais E., Thierry A.-M., Glowinski J. & Gioanni Y. Synaptic influence of hippocampus on pyramidal cells of the rat prefrontal cortex: an in vivo intracellular recording study. Cereb. Cortex N. Y. N 1991 13, 782–792 (2003). - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
