

Seeding and transgenic overexpression of alpha-synuclein triggers dendritic spine pathology in the neocortex
- PMID: 28351932
- PMCID: PMC5412764
- DOI: 10.15252/emmm.201607305
Seeding and transgenic overexpression of alpha-synuclein triggers dendritic spine pathology in the neocortex
Abstract
Although misfolded and aggregated α-synuclein (α-syn) is recognized in the disease progression of synucleinopathies, its role in the impairment of cortical circuitries and synaptic plasticity remains incompletely understood. We investigated how α-synuclein accumulation affects synaptic plasticity in the mouse somatosensory cortex using two distinct approaches. Long-term in vivo imaging of apical dendrites was performed in mice overexpressing wild-type human α-synuclein. Additionally, intracranial injection of preformed α-synuclein fibrils was performed to induce cortical α-syn pathology. We find that α-synuclein overexpressing mice show decreased spine density and abnormalities in spine dynamics in an age-dependent manner. We also provide evidence for the detrimental effects of seeded α-synuclein aggregates on dendritic architecture. We observed spine loss as well as dystrophic deformation of dendritic shafts in layer V pyramidal neurons. Our results provide a link to the pathophysiology underlying dementia associated with synucleinopathies and may enable the evaluation of potential drug candidates on dendritic spine pathology in vivo.
Keywords: alpha‐synuclein; dendritic spines; in vivo imaging; seeding; synucleinopathies.
© 2017 The Authors. Published under the terms of the CC BY 4.0 license.
Figures
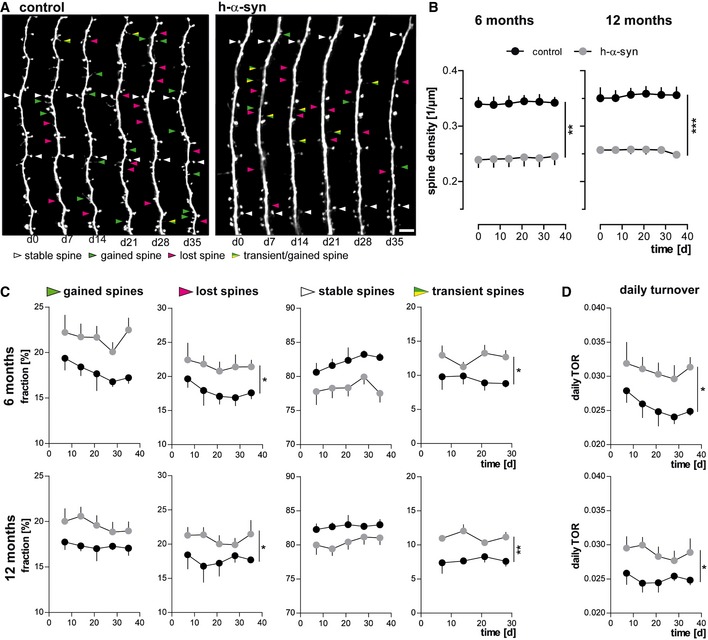
Representative in vivo two‐photon recordings of eGFP‐labeled apical tuft dendrites in the somatosensory cortex in h‐α‐syn and control animals. Arrowheads mark representative spines that were stable (white, present > 7 days), newly formed (green), or lost (magenta). Gained spines that do not stabilize (yellow/green, present < 7 days) are defined as transient. Scale bar, 5 μm.
Spine density is reduced in both 6‐ (**P = 0.0012) and 12‐month‐old (***P = 0.0001) h‐α‐syn animals.
The fractions of both gained (P 6 months = 0.0527; P 12 months = 0.0678) and lost spines (*P 6 months = 0.0461; *P 12 months = 0.0206) are elevated in h‐α‐syn mice compared to controls; the fraction of transient spines is significantly higher (*P 6 months = 0.0315; **P 12 months = 0.0017).
Consequently, the daily turnover ratio (TOR) is significantly increased in both 6‐ and 12‐month‐old h‐α‐syn mice (*P 6 months = 0.0491; *P 12 months = 0.0338).
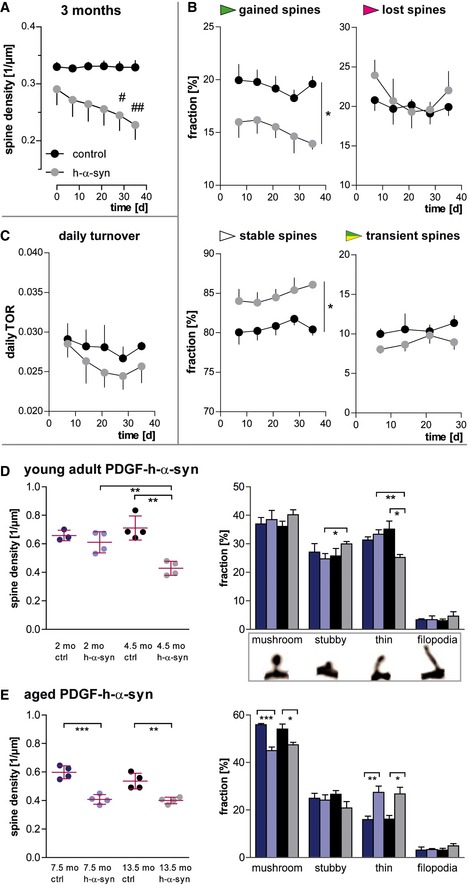
In young, 3‐month‐old h‐α‐syn mice, progressive decrease in spine density is observed in vivo.
The loss of spines in young h‐α‐syn animals is driven by a reduced fraction of newly gained spines, while the fraction of lost spines remains unchanged and the fraction of stable spines is increased (*P gained/stable = 0.0256).
The daily turnover of spines shows no significant difference between groups (P = 0.4062).
Ex vivo confocal data in young mice confirm synapse loss in h‐α‐syn mice between 2 and 4.5 months of age (**P syn 2/4.5 months = 0.0064; **P ctrl/syn 4.5 months = 0.0012) and show a shift in spine morphology toward relatively more stubby (*pstubby = 0.0418) and less thin spines (**P h‐α‐syn 2/4.5 months = 0.0044; *P ctrl/syn 4.5 months = 0.0155).
Ex vivo confocal data in aged h‐α‐syn mice show a decrease in total spine density (***P 7.5 months = 0.0005; **P 13.5 months = 0.0039) as well as in the fraction of mushroom spines (***P 7.5 months = 0.0004; *P 13.5 months = 0.0298), whereas the fraction of thin spines is increased (**P 7.5 months = 0.0097; *P 13.5 months = 0.0155).
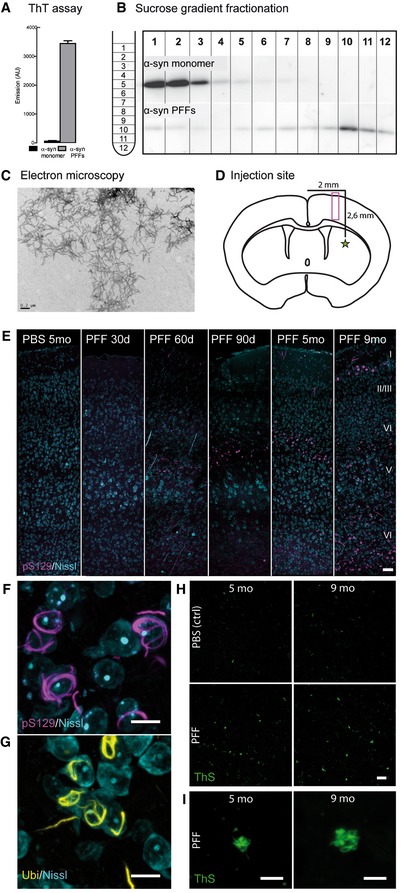
- A–C
The quality of the injection material was verified using ThT fluorescence assay (A), sucrose‐gradient fractionation (B), and electron microscopy (C). Scale bar: 0.2 μm.
- D
Injection site, 0.2 mm anterior of the bregma (star) and imaging area (box) as depicted in (E).
- E–I
Representative images of the cortical layers I–VI in the somatosensory cortex of mice at different time points postinjection. Controls were injected with sterile PBS. Neurons in the layers IV and V of the somatosensory cortex contain aggregates of α‐synuclein phosphorylated at S129 (F), which are ubiquitin‐positive (G) as well as thioflavin S‐positive (H, I). Image stacks (E–I) are depicted as maximum intensity projections. Scale bars: 50 μm (E), 10 μm (F, G), 20 μm (H), 5 μm (I).
α‐Synuclein aggregates occur as intrasomal (arrowheads) and neuritic (arrows) accumulations and are present predominantly in upper layer V and layer IV of the cortex. In layer I, pS129‐positive structures are much less dense. Lines exemplarily mark dendrites used for analysis.
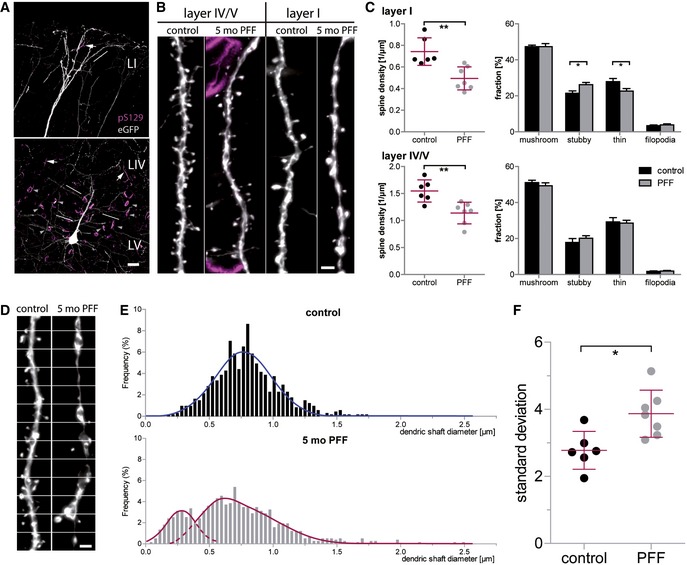
Spine analysis was performed on apical dendrites located in the cortical layers IV/V and I.
In layer I, spine density is reduced relative to PBS‐injected controls (**P = 0.0028), with the fraction of stubby spines being increased (*P = 0.0247) and the fraction of thin spines being decreased (*P = 0.0385). In layer IV/V, spine density is reduced as well (**P = 0.004), without a significant effect on spine morphology.
In apical tuft dendrites of PFF‐injected mice, dendrites display dystrophic swellings and parts of very small diameter; white lines: measurement positions.
Histograms of the dendritic shaft diameter.
Variation in the diameter of single dendrites in seeded mice compared to controls (*P = 0.011).
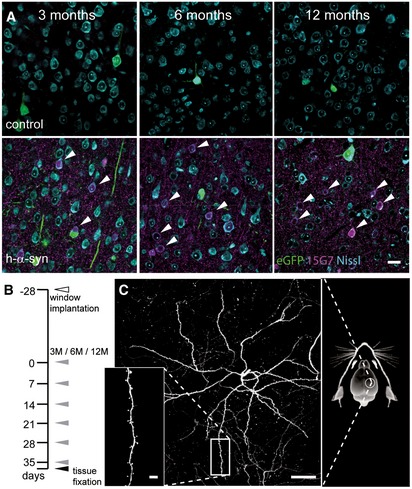
Immunostaining with 15G7 antibody shows cortical layer V overexpression of α‐synuclein (magenta) including accumulation in cell bodies (arrows).
Experimental timeline: 4 weeks after window implantation (white arrowhead), imaging was performed over 6 weeks, once every 7 days (gray arrowheads) and finally followed by perfusion and tissue fixation directly after the last imaging session (black arrowhead).
Overview and detailed projections of eGFP‐labeled layer V apical dendrites in the somatosensory cortex, imaged through a chronic cranial window.
References
-
- Aarsland D, Beyer MK, Kurz MW (2008) Dementia in Parkinson's disease. Curr Opin Neurol 21: 676–682 - PubMed
-
- Abeliovich A, Schmitz Y, Fariñas I, Choi‐Lundberg D, Ho W‐H, Castillo PE, Shinsky N, Verdugo JMG, Armanini M, Ryan A et al (2000) Mice lacking α‐Synuclein display functional deficits in the nigrostriatal dopamine system. Neuron 25: 239–252 - PubMed
-
- Arellano JI, Espinosa A, Fairén A, Yuste R, DeFelipe J (2007) Non‐synaptic dendritic spines in neocortex. Neuroscience 145: 464–469 - PubMed
-
- Bellucci A, Zaltieri M, Navarria L, Grigoletto J, Missale C, Spano P (2012) From α‐synuclein to synaptic dysfunctions: new insights into the pathophysiology of Parkinson's disease. Brain Res 1476: 183–202 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
