

Mitochondrial protein Fus1/Tusc2 in premature aging and age-related pathologies: critical roles of calcium and energy homeostasis
- PMID: 28351997
- PMCID: PMC5391223
- DOI: 10.18632/aging.101213
Mitochondrial protein Fus1/Tusc2 in premature aging and age-related pathologies: critical roles of calcium and energy homeostasis
Abstract
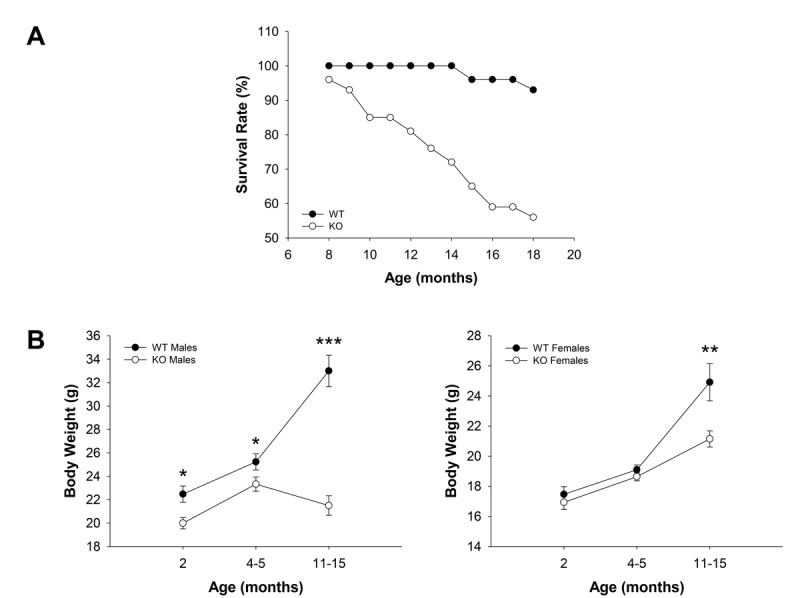
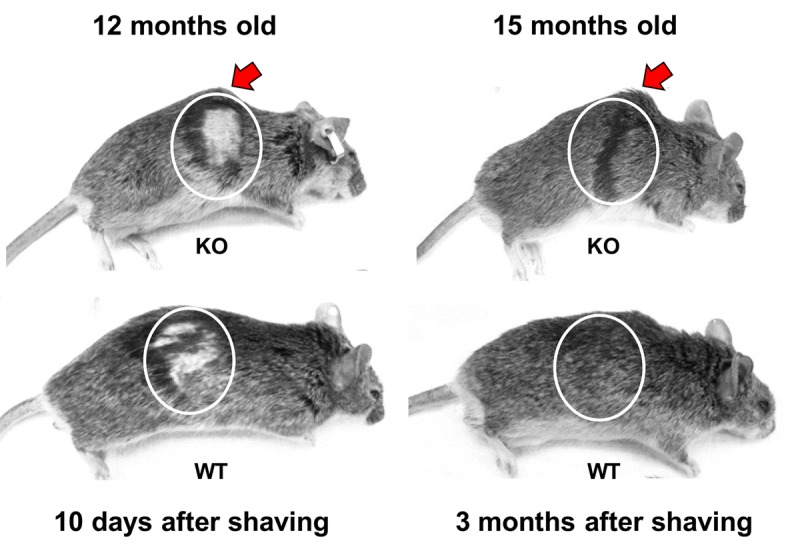
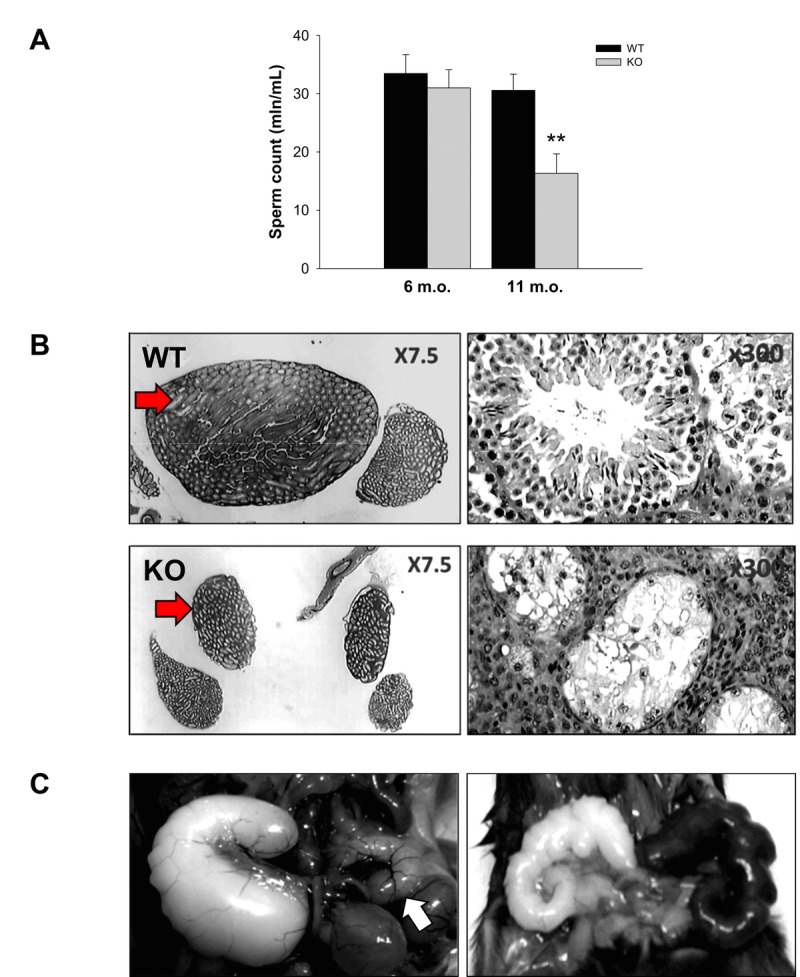
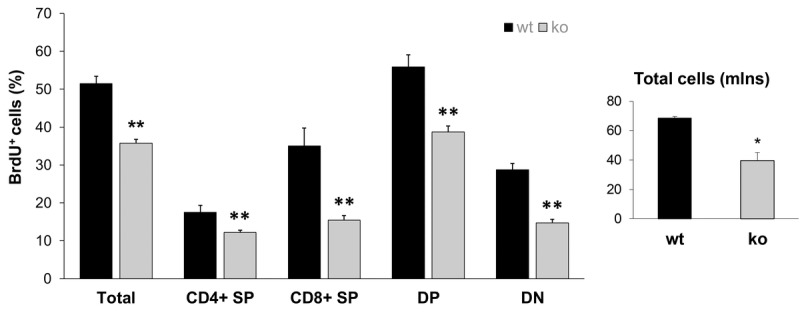
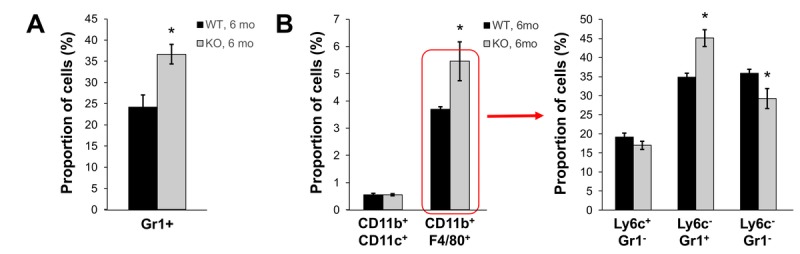
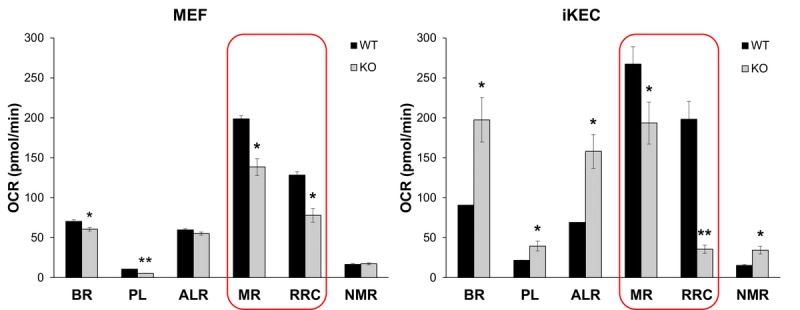
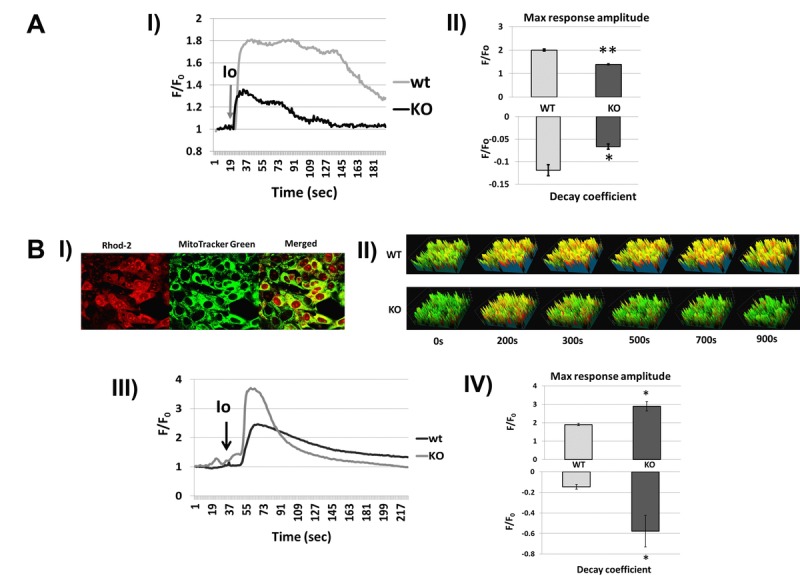
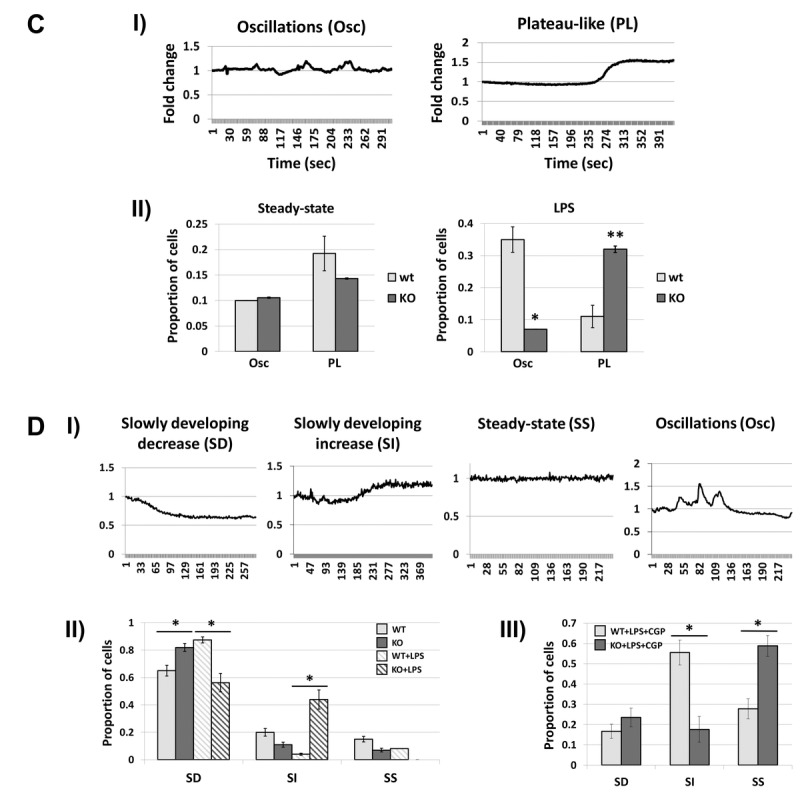
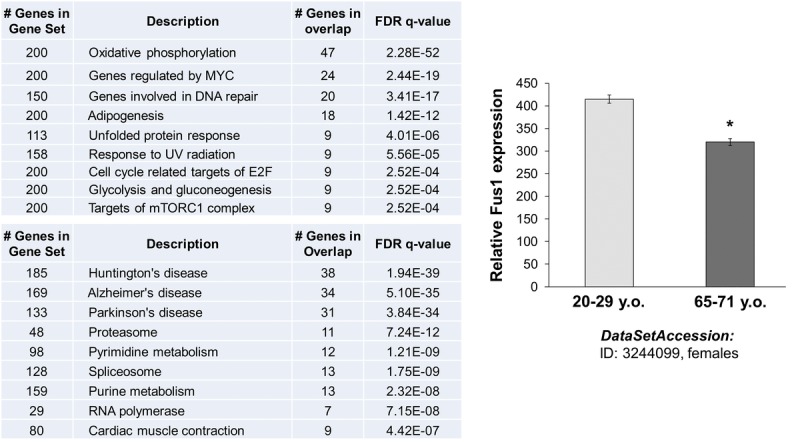
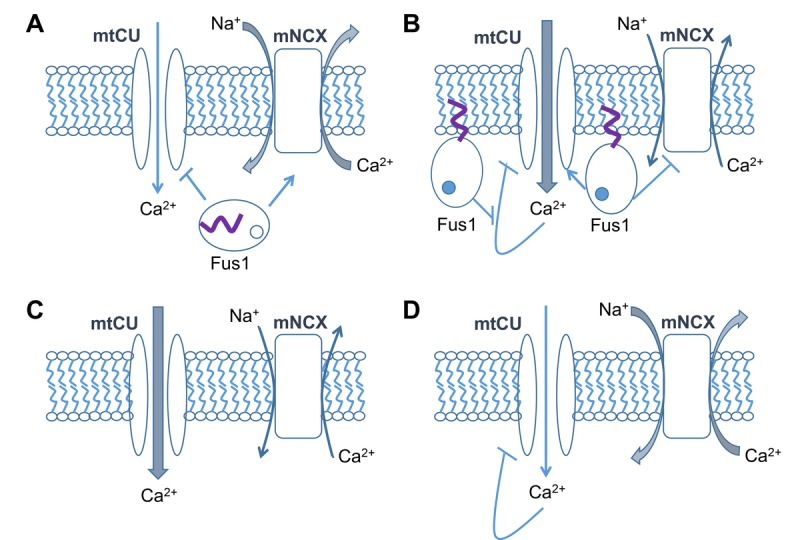
Decreased energy production and increased oxidative stress are considered to be major contributors to aging and aging-associated pathologies. The role of mitochondrial calcium homeostasis has also been highlighted as an important factor affecting different pathological conditions. Here, we present evidence that loss of a small mitochondrial protein Fus1 that maintains mitochondrial homeostasis results in premature aging, aging-associated pathologies, and decreased survival. We showed that Fus1KO mice develop multiple early aging signs including lordokyphosis, lack of vigor, inability to accumulate fat, reduced ability to tolerate stress, and premature death. Other prominent pathological changes included low sperm counts, compromised ability of adult stem cells to repopulate tissues, and chronic inflammation. At the molecular level, we demonstrated that mitochondria of Fus1 KO cells have low reserve respiratory capacity (the ability to produce extra energy during sudden energy demanding situations), and show significantly altered dynamics of cellular calcium response.Our recent studies on early hearing and memory loss in Fus1 KO mice combined with the new data presented here suggest that calcium and energy homeostasis controlled by Fus1 may be at the core of its aging-regulating activities. Thus, Fus1 protein and Fus1-dependent pathways and processes may represent new tools and targets for anti-aging strategies.
Keywords: Fus1/Tusc2; calcium response; chronic inflammation; mitochondrial Ca aging and age-related diseases 2+,; mitochondrial respiration.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
