

Membrane Oxidation in Cell Delivery and Cell Killing Applications
- PMID: 28355059
- PMCID: PMC5905413
- DOI: 10.1021/acschembio.7b00237
Membrane Oxidation in Cell Delivery and Cell Killing Applications
Abstract
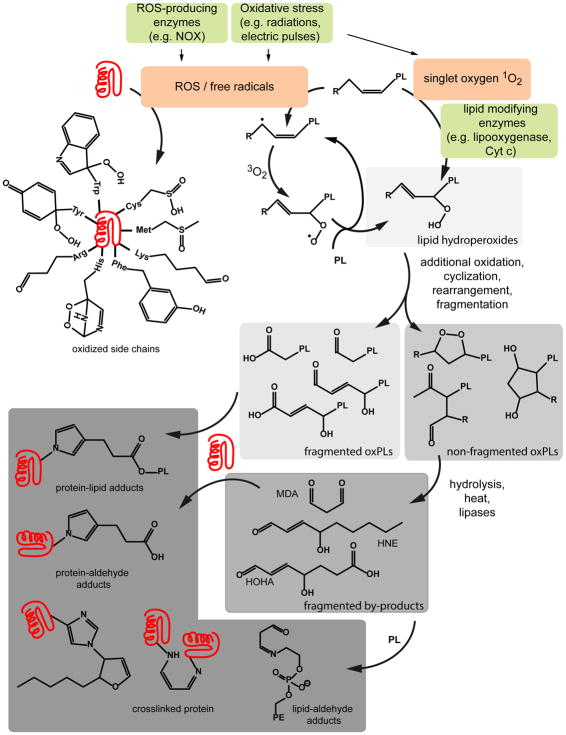
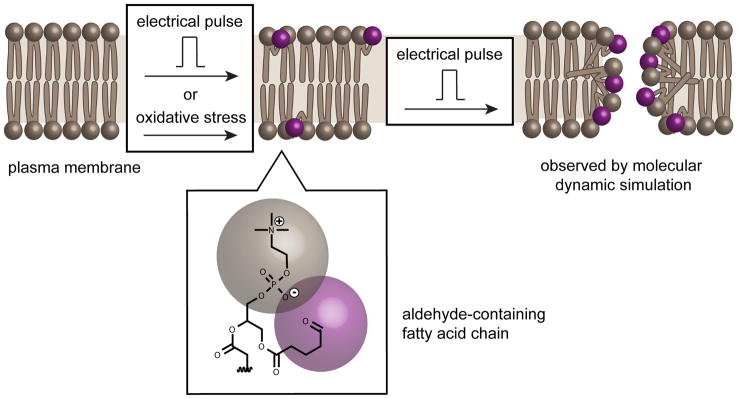
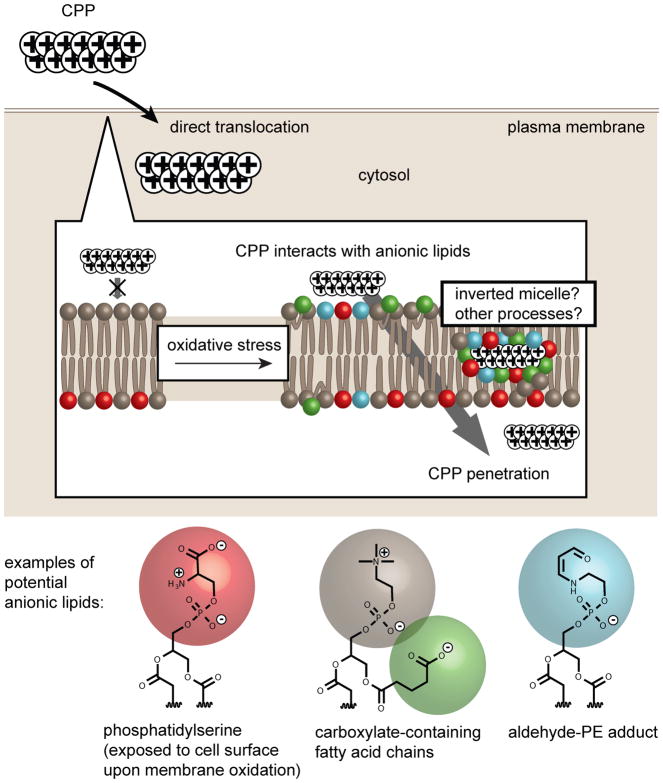
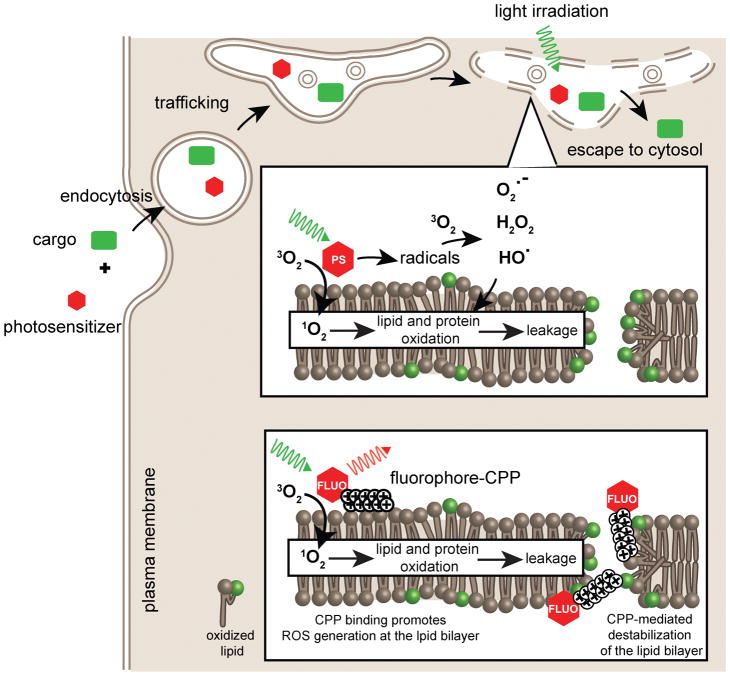
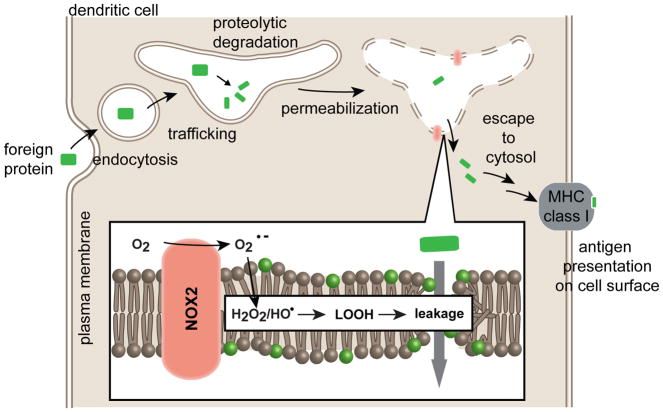
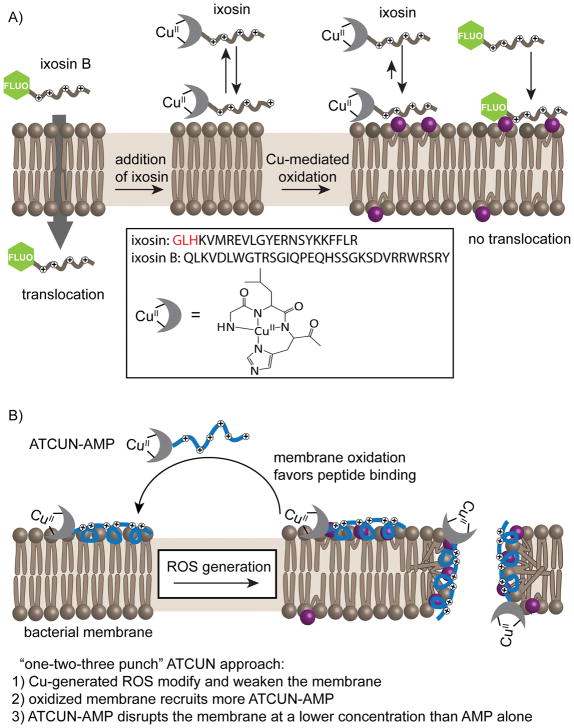
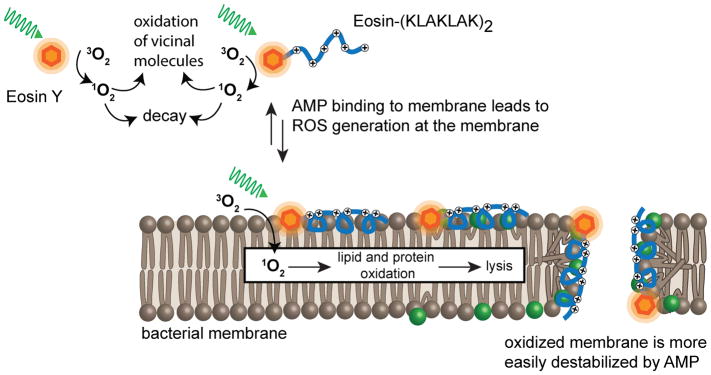
Cell delivery or cell killing processes often involve the crossing or disruption of cellular membranes. We review how, by modifying the composition and properties of membranes, membrane oxidation can be exploited to enhance the delivery of macromolecular cargoes into live human cells. We also describe how membrane oxidation can be utilized to achieve efficient killing of bacteria by antimicrobial peptides. Finally, we present recent evidence highlighting how membrane oxidation is intimately engaged in natural biological processes such as antigen delivery in dendritic cells and in the killing of bacteria by antimicrobial peptides. Overall, the insights that have been recently gained in this area should facilitate the development of more effective delivery technologies and antimicrobial therapeutic approaches.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Stewart MP, Sharei A, Ding X, Sahay G, Langer R, Jensen KF. In vitro and ex vivo strategies for intracellular delivery. Nature. 2016;538:183–192. - PubMed
-
- Lewis K. Platforms for antibiotic discovery. Nat Rev Drug Discov. 2013;12:371–387. - PubMed
-
- Halliwell B. Cell culture, oxidative stress, and antioxidants: avoiding pitfalls. Biomedical journal. 2014;37:99–105. - PubMed
-
- Reis A, Spickett CM. Chemistry of phospholipid oxidation. Biochimica et biophysica acta. 2012;1818:2374–2387. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
