

Volume Transitions of Isolated Cell Nuclei Induced by Rapid Temperature Increase
- PMID: 28355535
- PMCID: PMC5374986
- DOI: 10.1016/j.bpj.2017.01.022
Volume Transitions of Isolated Cell Nuclei Induced by Rapid Temperature Increase
Abstract
Understanding the physical mechanisms governing nuclear mechanics is important as it can impact gene expression and development. However, how cell nuclei respond to external cues such as heat is not well understood. Here, we studied the material properties of isolated nuclei in suspension using an optical stretcher. We demonstrate that isolated nuclei regulate their volume in a highly temperature-sensitive manner. At constant temperature, isolated nuclei behaved like passive, elastic and incompressible objects, whose volume depended on the pH and ionic conditions. When the temperature was increased suddenly by even a few degrees Kelvin, nuclei displayed a repeatable and reversible temperature-induced volume transition, whose sign depended on the valency of the solvent. Such phenomenon is not observed for nuclei subjected to slow heating. The transition temperature could be shifted by adiabatic changes of the ambient temperature, and the magnitude of temperature-induced volume transition could be modulated by modifying the chromatin compaction state and remodeling processes. Our findings reveal that the cell nucleus can be viewed as a highly charged polymer gel with intriguing thermoresponsive properties, which might play a role in nuclear volume regulation and thermosensing in living cells.
Copyright © 2017 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
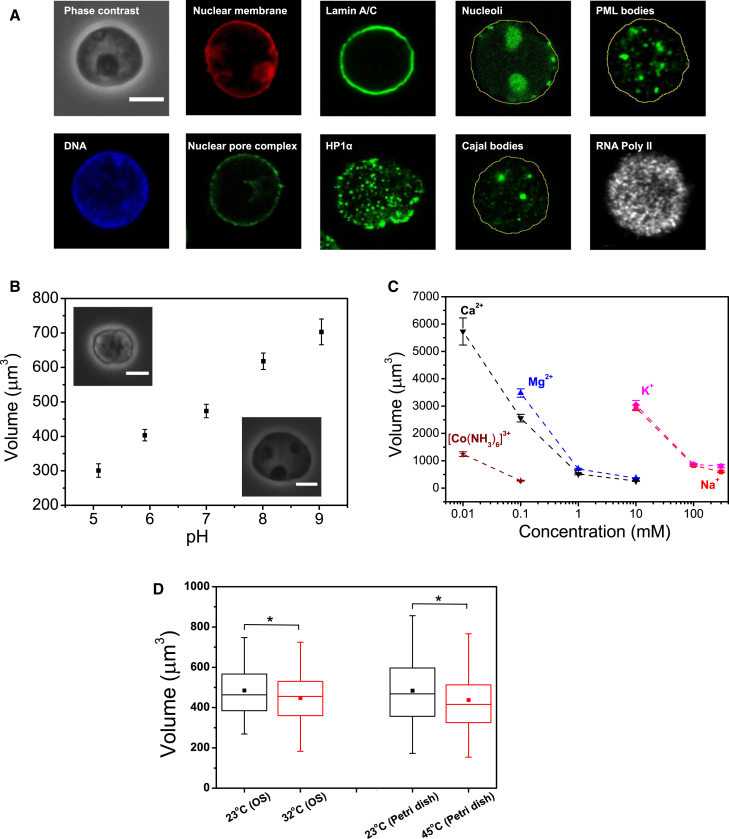
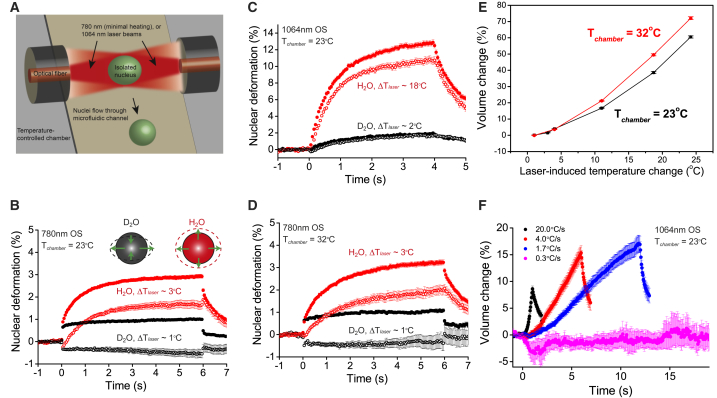
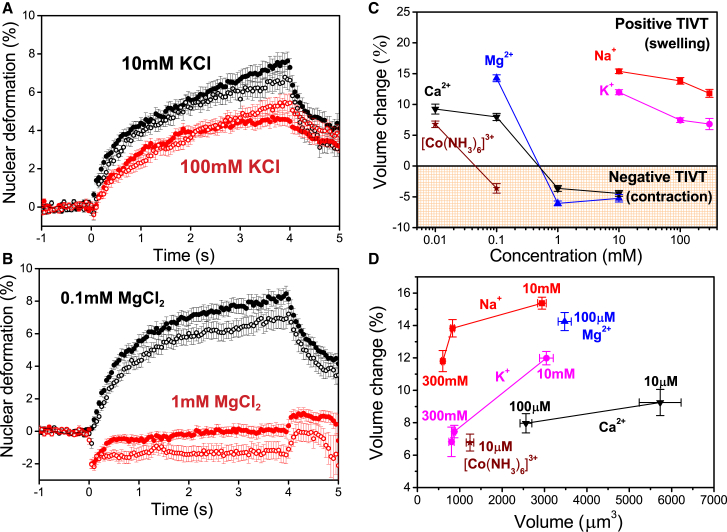
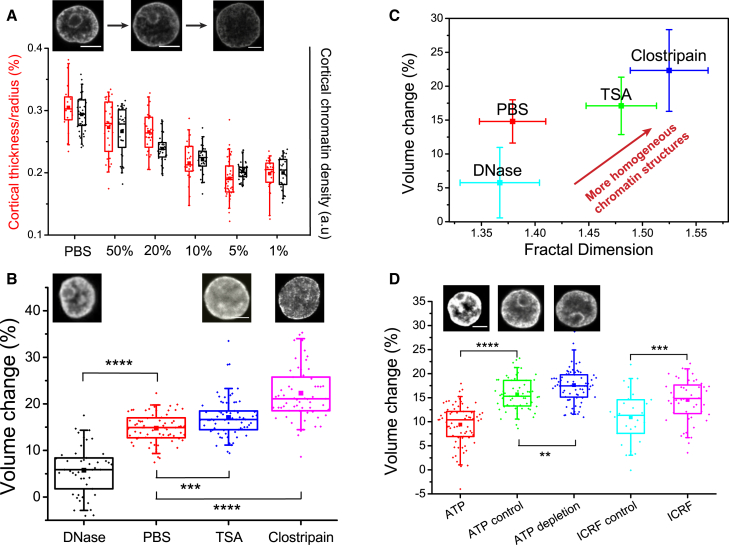
References
-
- Guilak F., Tedrow J.R., Burgkart R. Viscoelastic properties of the cell nucleus. Biochem. Biophys. Res. Commun. 2000;269:781–786. - PubMed
-
- Versaevel M., Grevesse T., Gabriele S. Spatial coordination between cell and nuclear shape within micropatterned endothelial cells. Nat. Commun. 2012;3:671. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
