

Hydrodynamic Hunters
- PMID: 28355554
- PMCID: PMC5376100
- DOI: 10.1016/j.bpj.2017.02.011
Hydrodynamic Hunters
Abstract
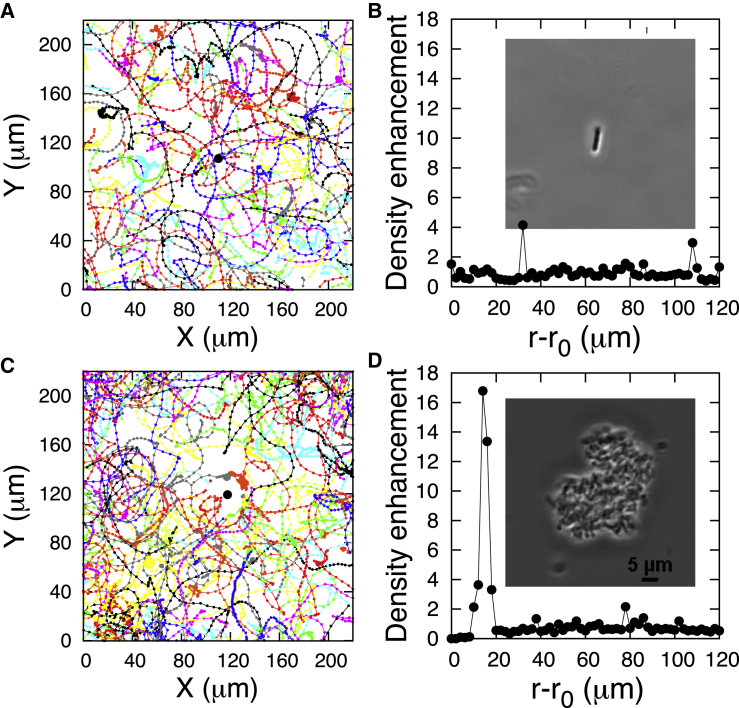
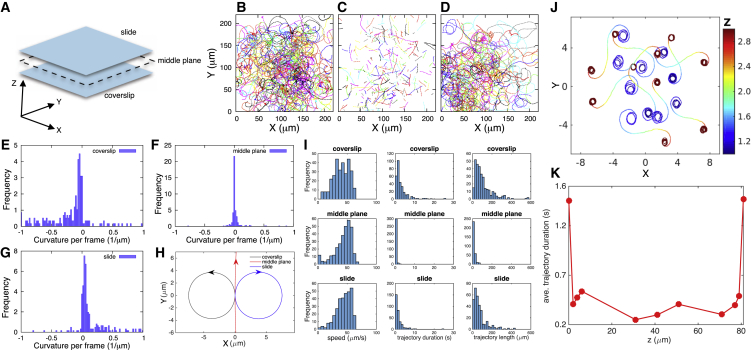
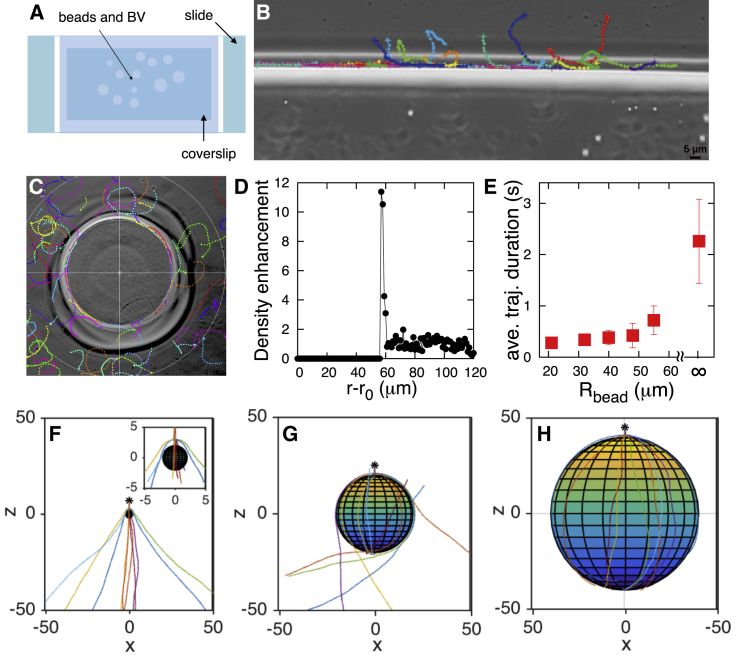
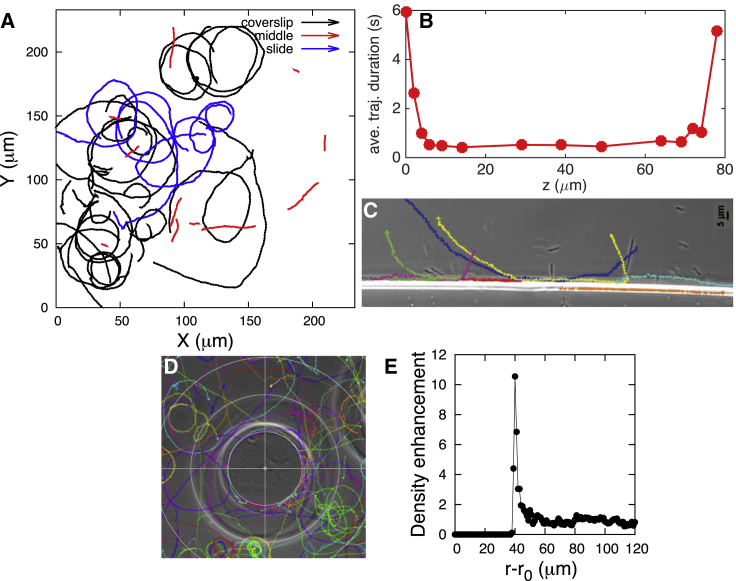
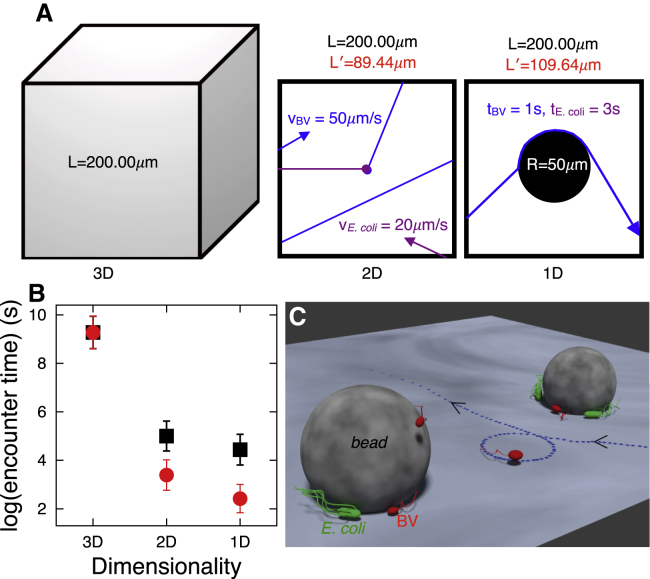
The Gram-negative Bdellovibrio bacteriovorus (BV) is a model bacterial predator that hunts other bacteria and may serve as a living antibiotic. Despite over 50 years since its discovery, it is suggested that BV probably collides into its prey at random. It remains unclear to what degree, if any, BV uses chemical cues to target its prey. The targeted search problem by the predator for its prey in three dimensions is a difficult problem: it requires the predator to sensitively detect prey and forecast its mobile prey's future position on the basis of previously detected signal. Here instead we find that rather than chemically detecting prey, hydrodynamics forces BV into regions high in prey density, thereby improving its odds of a chance collision with prey and ultimately reducing BV's search space for prey. We do so by showing that BV's dynamics are strongly influenced by self-generated hydrodynamic flow fields forcing BV onto surfaces and, for large enough defects on surfaces, forcing BV in orbital motion around these defects. Key experimental controls and calculations recapitulate the hydrodynamic origin of these behaviors. While BV's prey (Escherichia coli) are too small to trap BV in hydrodynamic orbit, the prey are also susceptible to their own hydrodynamic fields, substantially confining them to surfaces and defects where mobile predator and prey density is now dramatically enhanced. Colocalization, driven by hydrodynamics, ultimately reduces BV's search space for prey from three to two dimensions (on surfaces) even down to a single dimension (around defects). We conclude that BV's search for individual prey remains random, as suggested in the literature, but confined, however-by generic hydrodynamic forces-to reduced dimensionality.
Copyright © 2017 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
If Life Keeps Throwing Curveballs, You've Probably Reached a Wall.Biophys J. 2017 Mar 28;112(6):1047-1049. doi: 10.1016/j.bpj.2016.12.053. Biophys J. 2017. PMID: 28355533 Free PMC article. No abstract available.
References
-
- Stolp H., Starr M.P. Bdellovibrio bacteriovorus gen. et sp. n., a predatory, ectoparasitic, and bacteriolytic microorganism. Antonie van Leeuwenhoek. 1963;29:217–248. - PubMed
-
- Rendulic S., Jagtap P., Schuster S.C. A predator unmasked: life cycle of Bdellovibrio bacteriovorus from a genomic perspective. Science. 2004;303:689–692. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
