

Enhanced Degradation of Misfolded Proteins Promotes Tumorigenesis
- PMID: 28355566
- PMCID: PMC5603913
- DOI: 10.1016/j.celrep.2017.03.010
Enhanced Degradation of Misfolded Proteins Promotes Tumorigenesis
Abstract
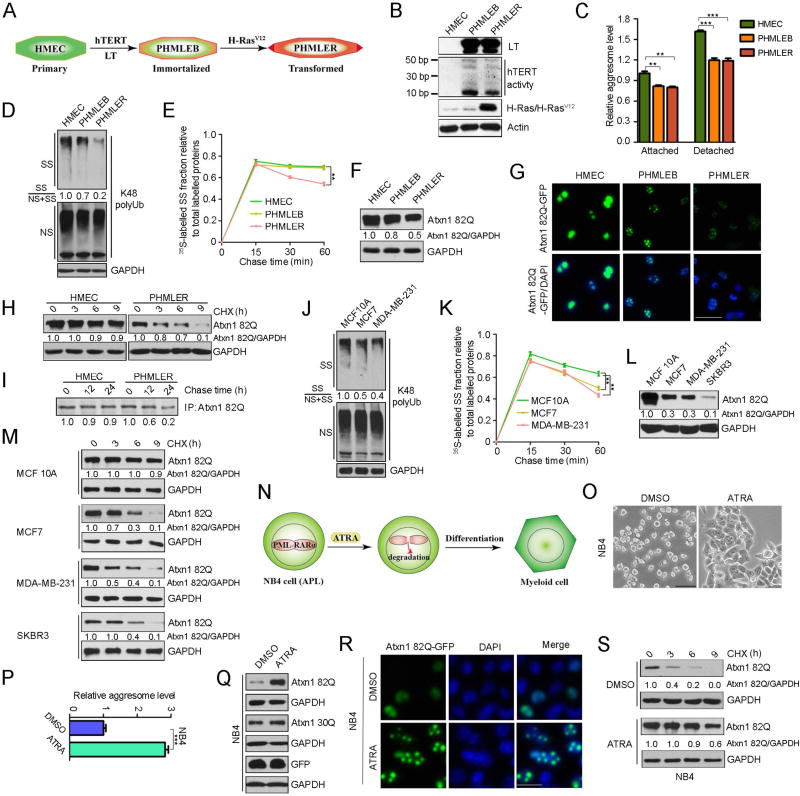
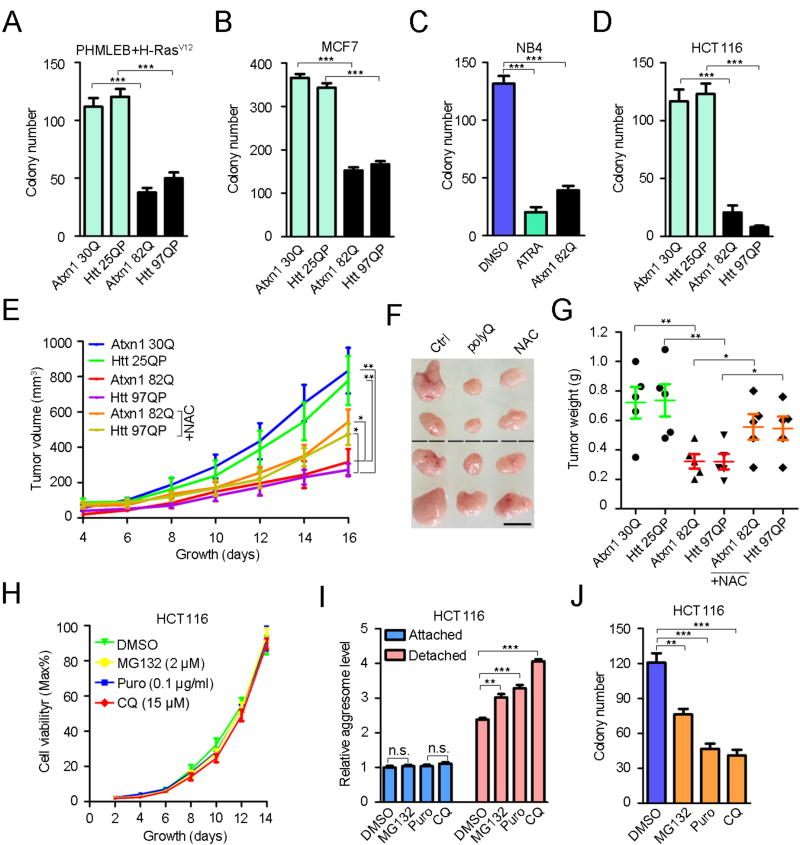
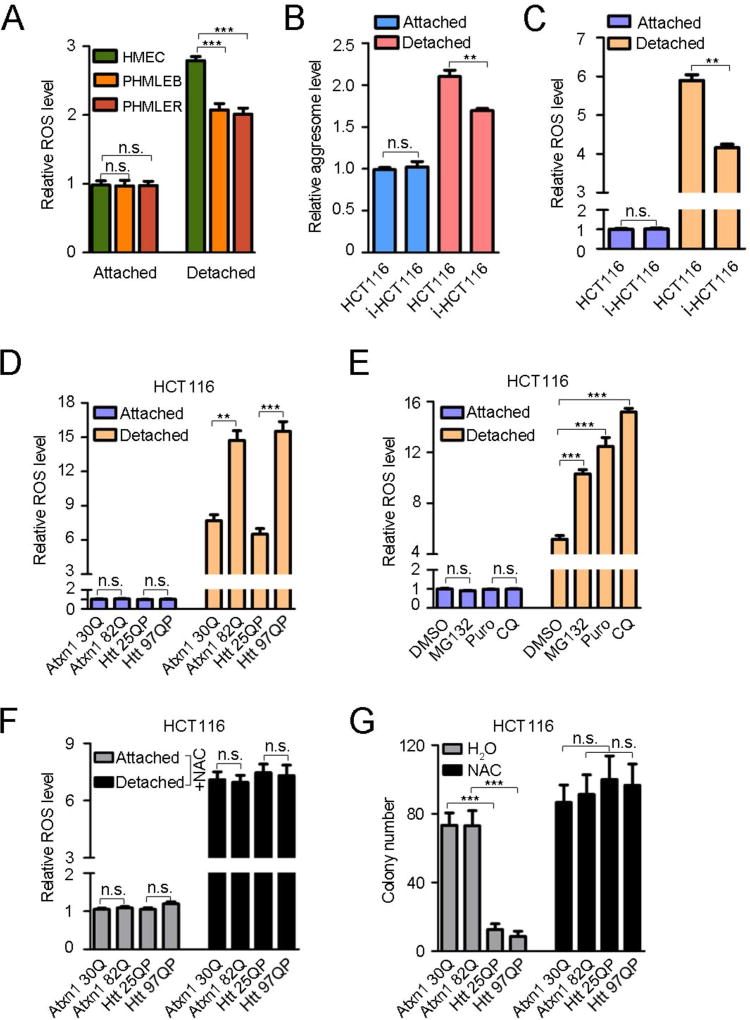
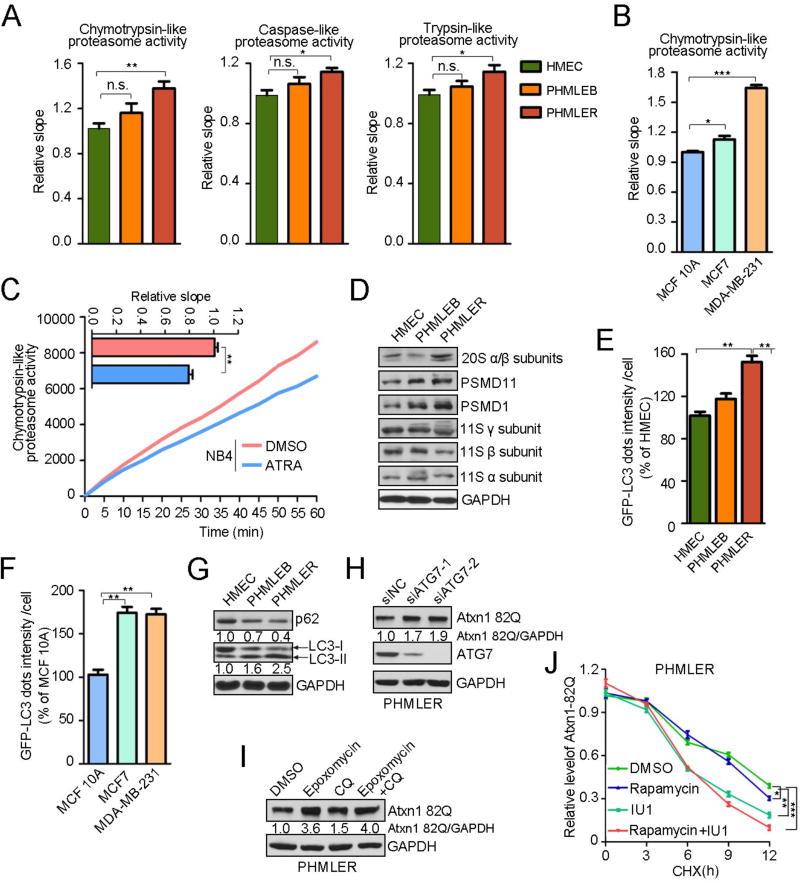
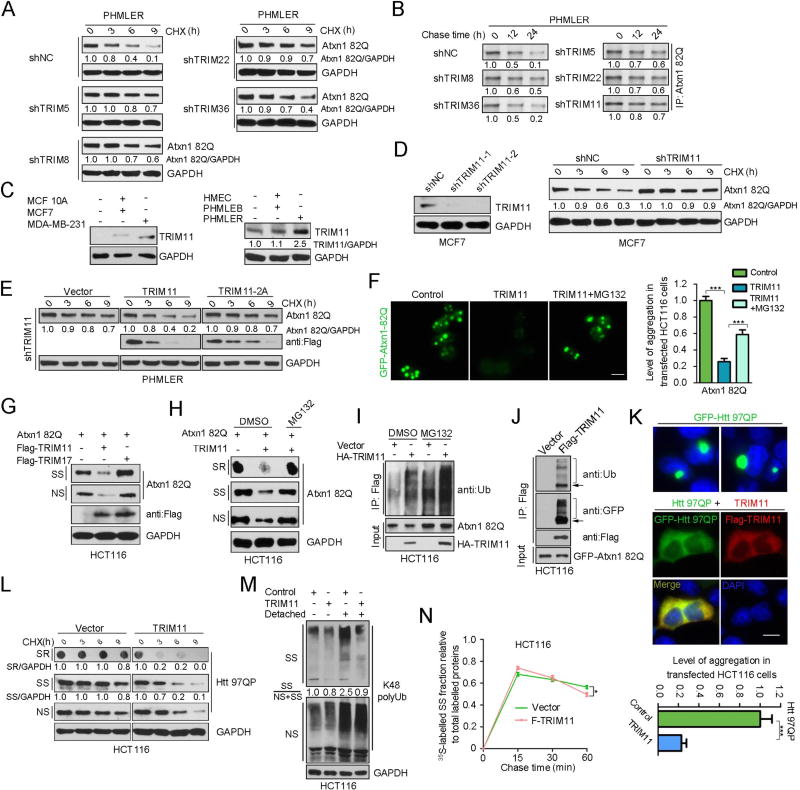
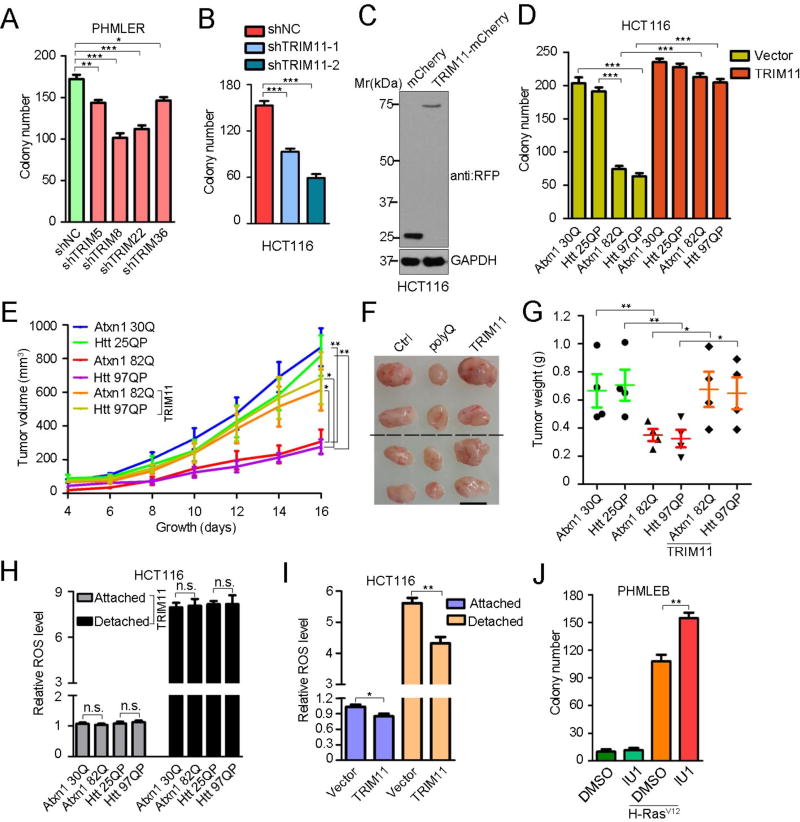
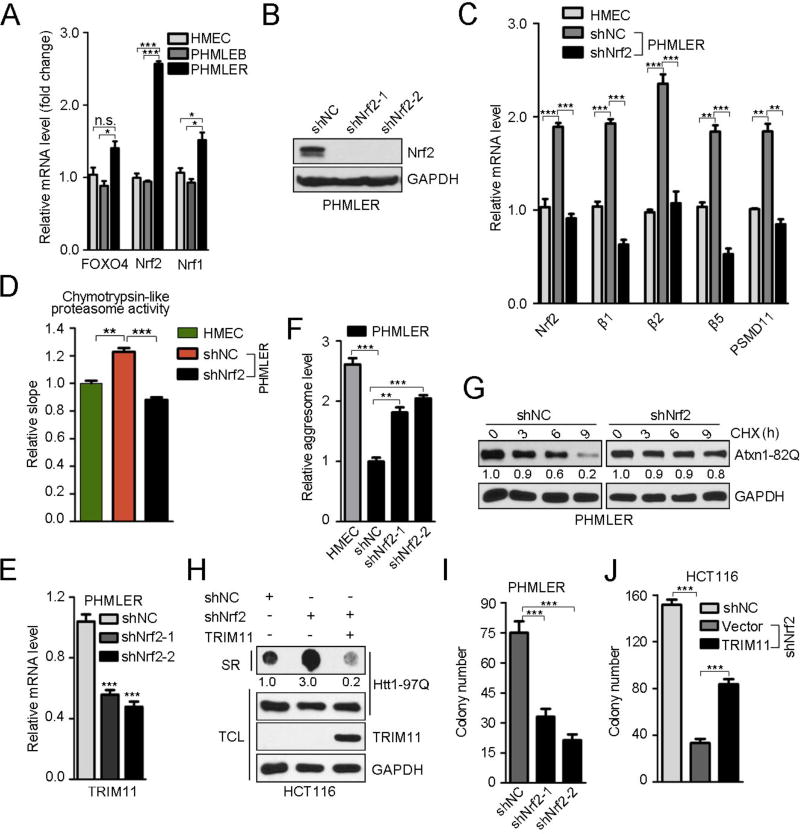
An adequate cellular capacity to degrade misfolded proteins is critical for cell survival and organismal health. A diminished capacity is associated with aging and neurodegenerative diseases; however, the consequences of an enhanced capacity remain undefined. Here, we report that the ability to clear misfolded proteins is increased during oncogenic transformation and is reduced upon tumor cell differentiation. The augmented capacity mitigates oxidative stress associated with oncogenic growth and is required for both the initiation and maintenance of malignant phenotypes. We show that tripartite motif-containing (TRIM) proteins select misfolded proteins for proteasomal degradation. The higher degradation power in tumor cells is attributed to the upregulation of the proteasome and especially TRIM proteins, both mediated by the antioxidant transcription factor Nrf2. These findings establish a critical role of TRIMs in protein quality control, connect the clearance of misfolded proteins to antioxidant defense, and suggest an intrinsic characteristic of tumor cells.
Keywords: Nrf2; PML; ROS; TRIM proteins; TRIM11; breast cancer; misfolded proteins; proteasome; protein quality control; transformation.
Copyright © 2017 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Arlt A, Bauer I, Schafmayer C, Tepel J, Muerkoster SS, Brosch M, Roder C, Kalthoff H, Hampe J, Moyer MP, et al. Increased proteasome subunit protein expression and proteasome activity in colon cancer relate to an enhanced activation of nuclear factor E2-related factor 2 (Nrf2) Oncogene. 2009;28:3983–3996. - PubMed
-
- Balch WE, Morimoto RI, Dillin A, Kelly JW. Adapting proteostasis for disease intervention. Science. 2008;319:916–919. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
