

Origins and Evolution of Stomatal Development
- PMID: 28356502
- PMCID: PMC5462063
- DOI: 10.1104/pp.17.00183
Origins and Evolution of Stomatal Development
Abstract
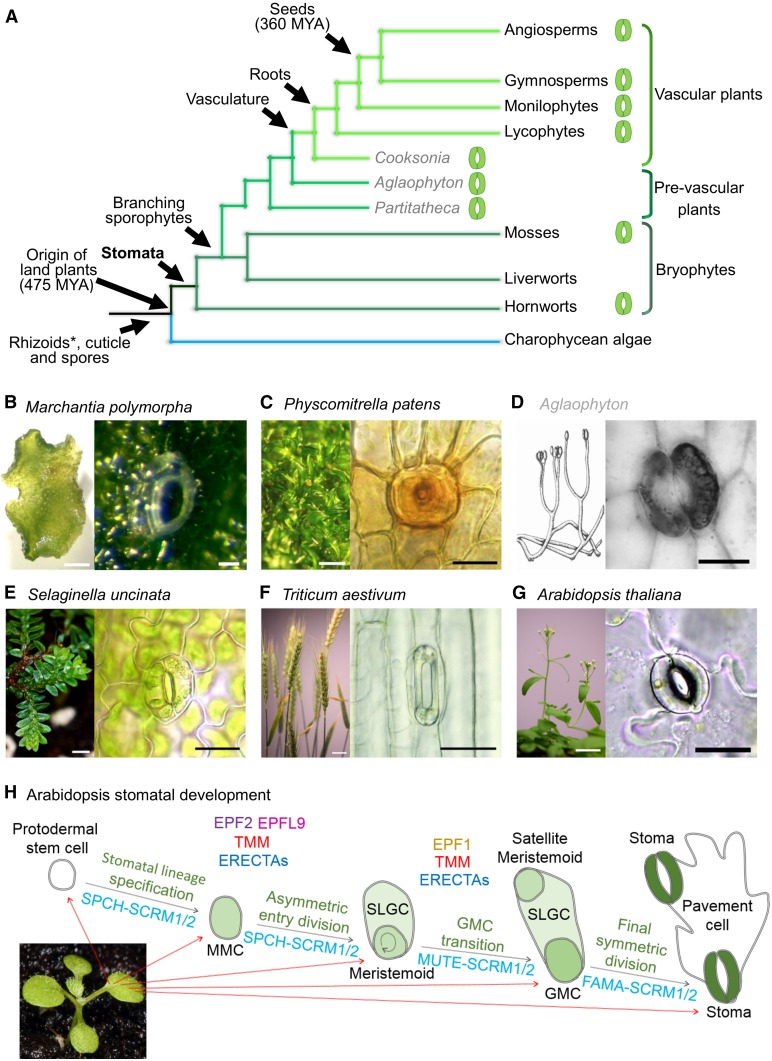
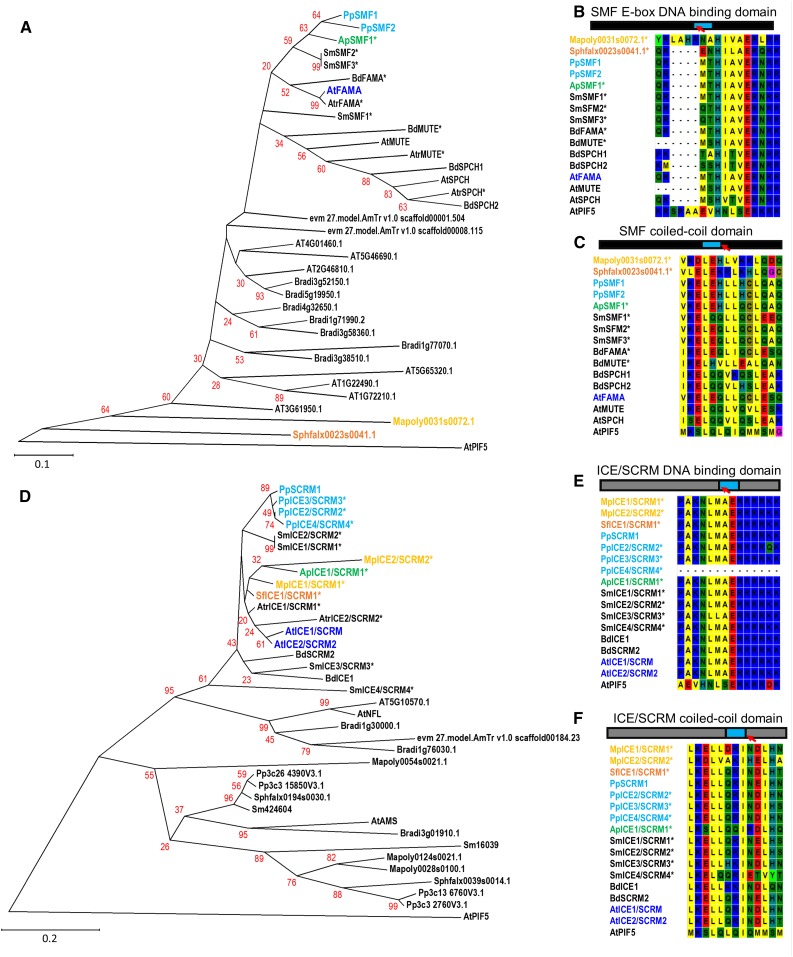
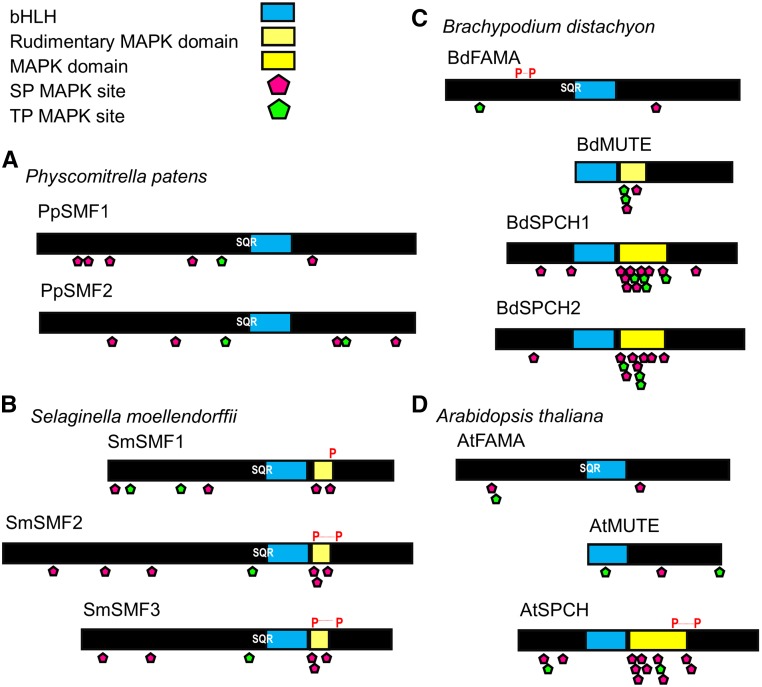
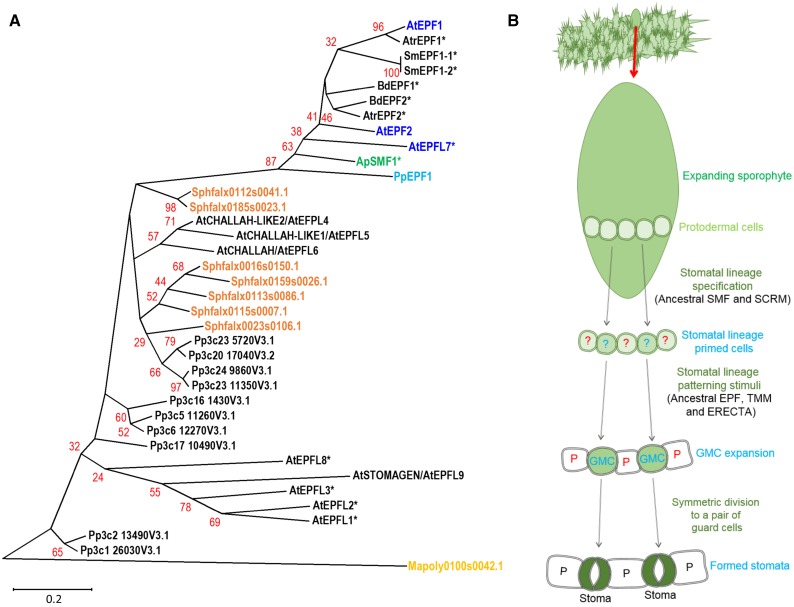
The fossil record suggests stomata-like pores were present on the surfaces of land plants over 400 million years ago. Whether stomata arose once or whether they arose independently across newly evolving land plant lineages has long been a matter of debate. In Arabidopsis, a genetic toolbox has been identified that tightly controls stomatal development and patterning. This includes the basic helix-loop-helix (bHLH) transcription factors SPEECHLESS (SPCH), MUTE, FAMA, and ICE/SCREAMs (SCRMs), which promote stomatal formation. These factors are regulated via a signaling cascade, which includes mobile EPIDERMAL PATTERNING FACTOR (EPF) peptides to enforce stomatal spacing. Mosses and hornworts, the most ancient extant lineages to possess stomata, possess orthologs of these Arabidopsis (Arabidopsis thaliana) stomatal toolbox genes, and manipulation in the model bryophyte Physcomitrella patens has shown that the bHLH and EPF components are also required for moss stomatal development and patterning. This supports an ancient and tightly conserved genetic origin of stomata. Here, we review recent discoveries and, by interrogating newly available plant genomes, we advance the story of stomatal development and patterning across land plant evolution. Furthermore, we identify potential orthologs of the key toolbox genes in a hornwort, further supporting a single ancient genetic origin of stomata in the ancestor to all stomatous land plants.
© 2017 American Society of Plant Biologists. All Rights Reserved.
Figures


References
-
- Assmann SM, Jegla T (2016) Guard cell sensory systems: Recent insights on stomatal responses to light, abscisic acid, and CO2. Curr Opin Plant Biol 33: 157–167 - PubMed
-
- Baars C, Edwards D (2008) Effects of elevated atmospheric CO2 on spore capsules of the moss Leptobryum pyriforme. J Bryol 30: 36–40
-
- Berry JA, Beerling DJ, Franks PJ (2010) Stomata: Key players in the earth system, past and present. Curr Opin Plant Biol 13: 233–240 - PubMed
-
- Bowman JL. (2011) Stomata: Active portals for flourishing on land. Curr Biol 21: R540–R541 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
