

Mutual Interplay between the Human Cytomegalovirus Terminase Subunits pUL51, pUL56, and pUL89 Promotes Terminase Complex Formation
- PMID: 28356534
- PMCID: PMC5446633
- DOI: 10.1128/JVI.02384-16
Mutual Interplay between the Human Cytomegalovirus Terminase Subunits pUL51, pUL56, and pUL89 Promotes Terminase Complex Formation
Abstract
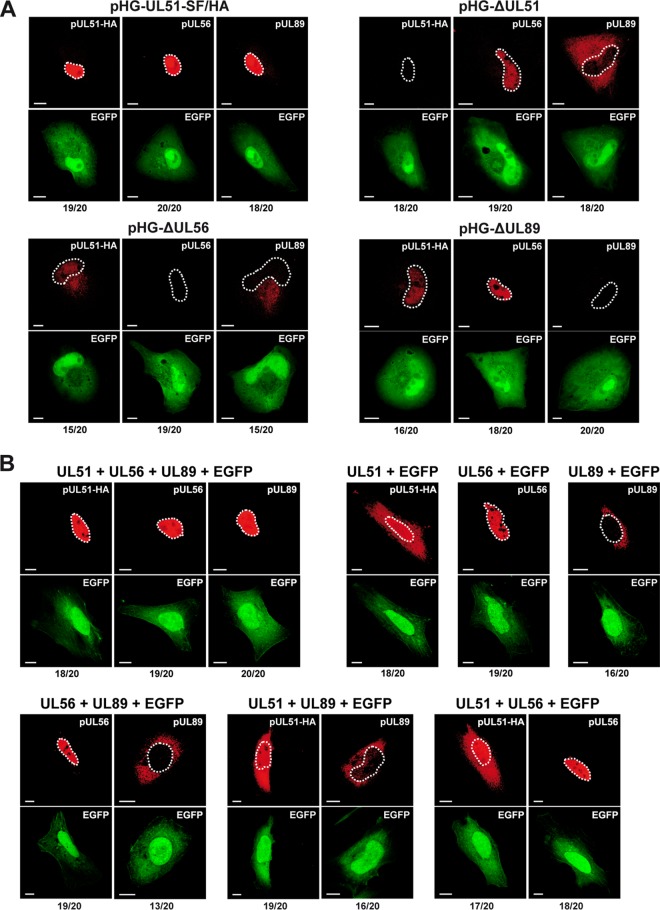
Human cytomegalovirus (HCMV) genome encapsidation requires several essential viral proteins, among them pUL56, pUL89, and the recently described pUL51, which constitute the viral terminase. To gain insight into terminase complex assembly, we investigated interactions between the individual subunits. For analysis in the viral context, HCMV bacterial artificial chromosomes carrying deletions in the open reading frames encoding the terminase proteins were used. These experiments were complemented by transient-transfection assays with plasmids expressing the terminase components. We found that if one terminase protein was missing, the levels of the other terminase proteins were markedly diminished, which could be overcome by proteasome inhibition or providing the missing subunit in trans These data imply that sequestration of the individual subunits within the terminase complex protects them from proteasomal turnover. The finding that efficient interactions among the terminase proteins occurred only when all three were present together is reminiscent of a folding-upon-binding principle leading to cooperative stability. Furthermore, whereas pUL56 was translocated into the nucleus on its own, correct nuclear localization of pUL51 and pUL89 again required all three terminase constituents. Altogether, these features point to a model of the HCMV terminase as a multiprotein complex in which the three players regulate each other concerning stability, subcellular localization, and assembly into the functional tripartite holoenzyme.IMPORTANCE HCMV is a major risk factor in immunocompromised individuals, and congenital CMV infection is the leading viral cause for long-term sequelae, including deafness and mental retardation. The current treatment of CMV disease is based on drugs sharing the same mechanism, namely, inhibiting viral DNA replication, and often results in adverse side effects and the appearance of resistant virus strains. Recently, the HCMV terminase has emerged as an auspicious target for novel antiviral drugs. A new drug candidate inhibiting the HCMV terminase, Letermovir, displayed excellent potency in clinical trials; however, its precise mode of action is not understood yet. Here, we describe the mutual dependence of the HCMV terminase constituents for their assembly into a functional terminase complex. Besides providing new basic insights into terminase formation, these results will be valuable when studying the mechanism of action for drugs targeting the HCMV terminase and developing additional substances interfering with viral genome encapsidation.
Keywords: complex assembly; cytomegalovirus; nuclear import; protein interactions; protein stability; terminase.
Copyright © 2017 American Society for Microbiology.
Figures
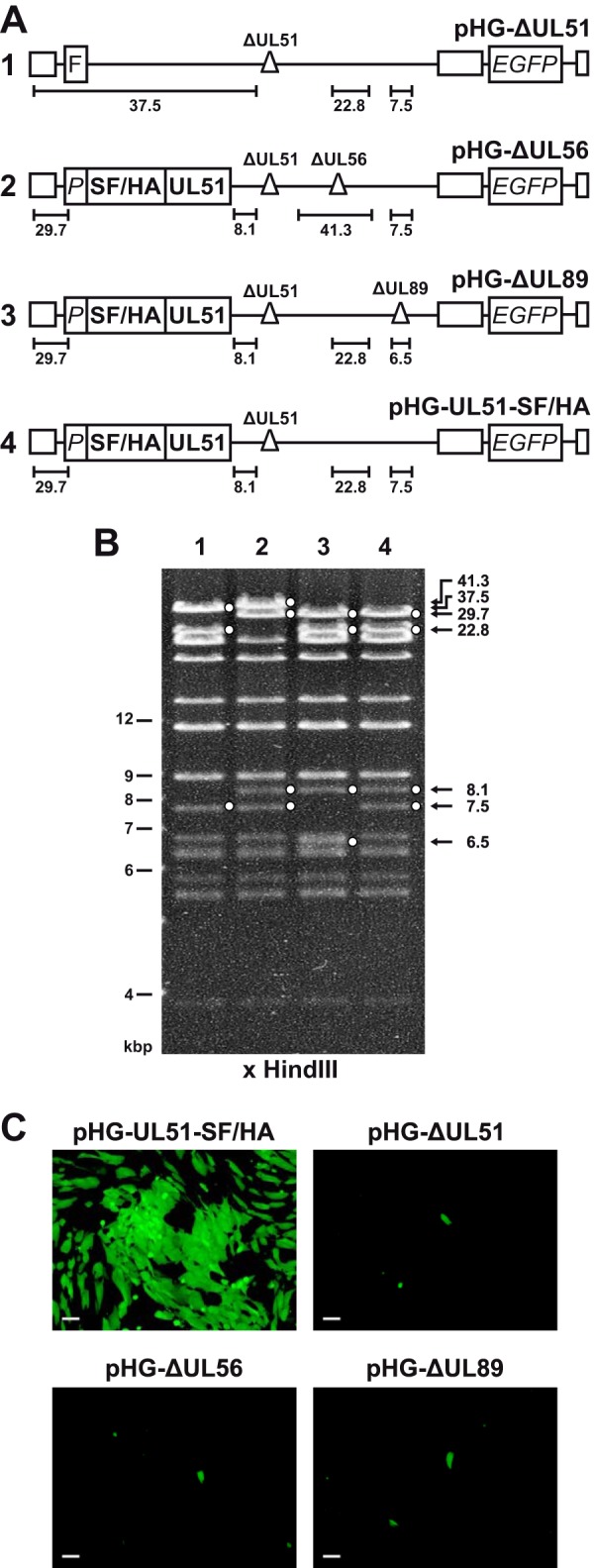
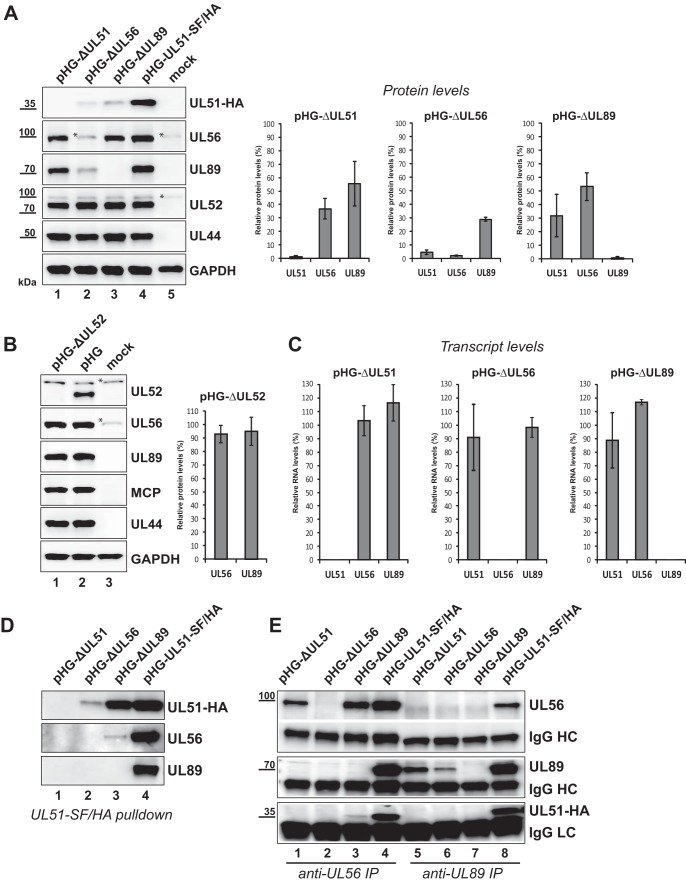
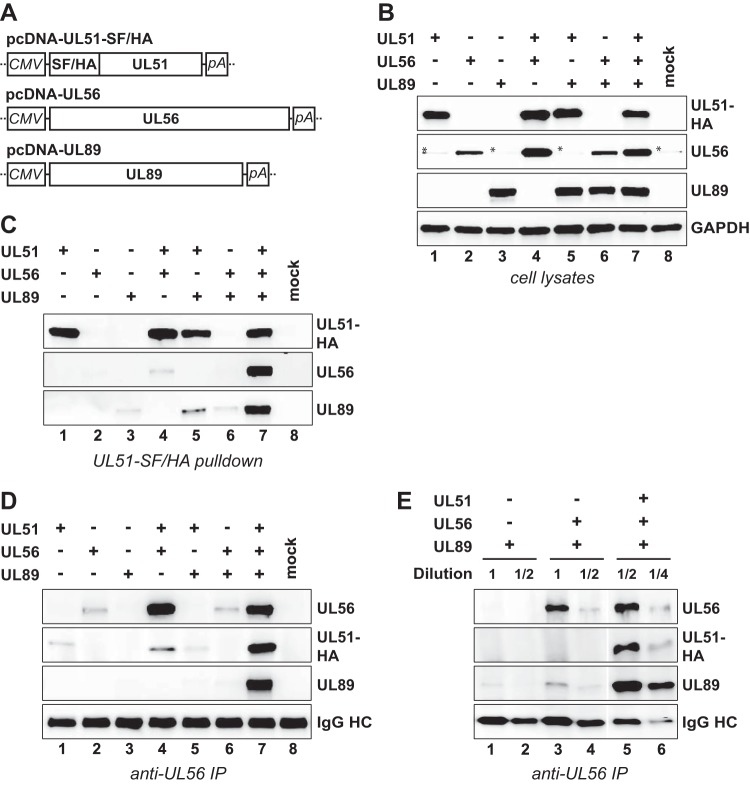
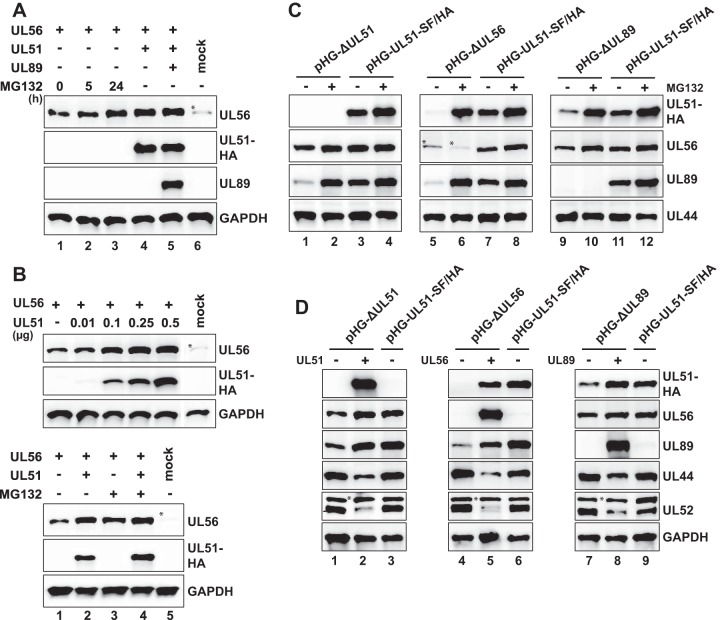
References
-
- Dittmer A, Drach JC, Townsend LB, Fischer A, Bogner E. 2005. Interaction of the putative human cytomegalovirus portal protein pUL104 with the large terminase subunit pUL56 and its inhibition by benzimidazole-d-ribonucleosides. J Virol 79:14660–14667. doi: 10.1128/JVI.79.23.14660-14667.2005. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
