

Herpes Simplex Virus 1 UL34 Protein Regulates the Global Architecture of the Endoplasmic Reticulum in Infected Cells
- PMID: 28356536
- PMCID: PMC5446647
- DOI: 10.1128/JVI.00271-17
Herpes Simplex Virus 1 UL34 Protein Regulates the Global Architecture of the Endoplasmic Reticulum in Infected Cells
Abstract
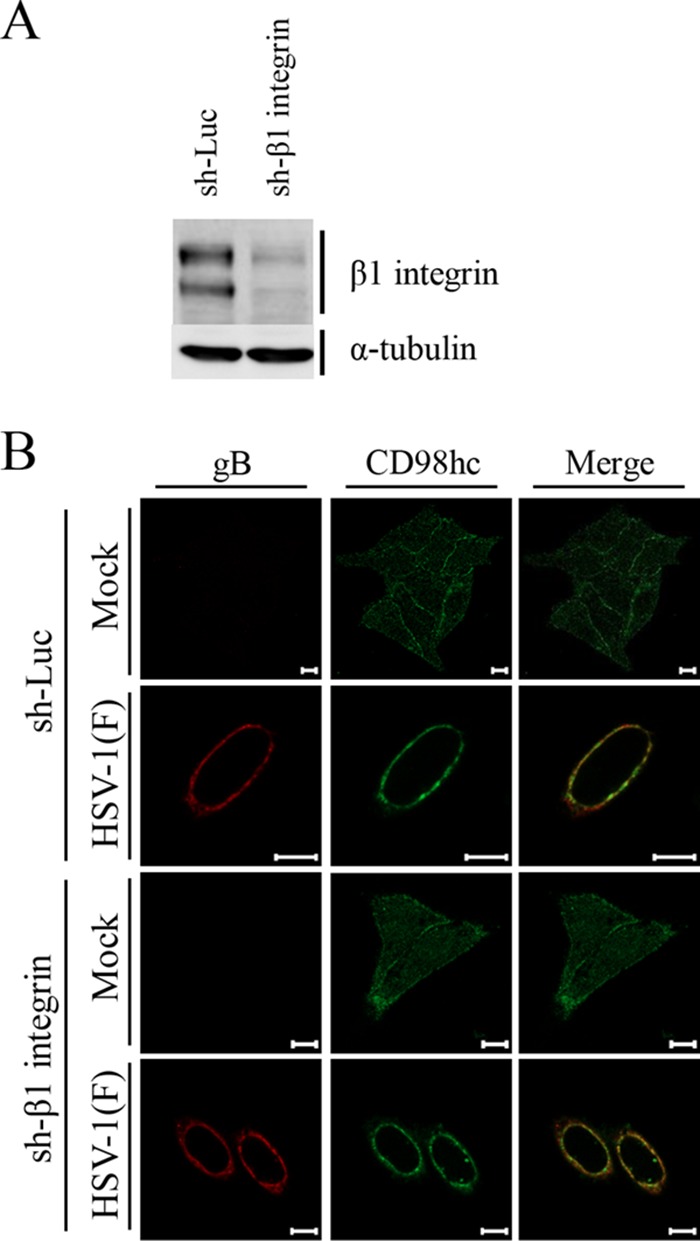
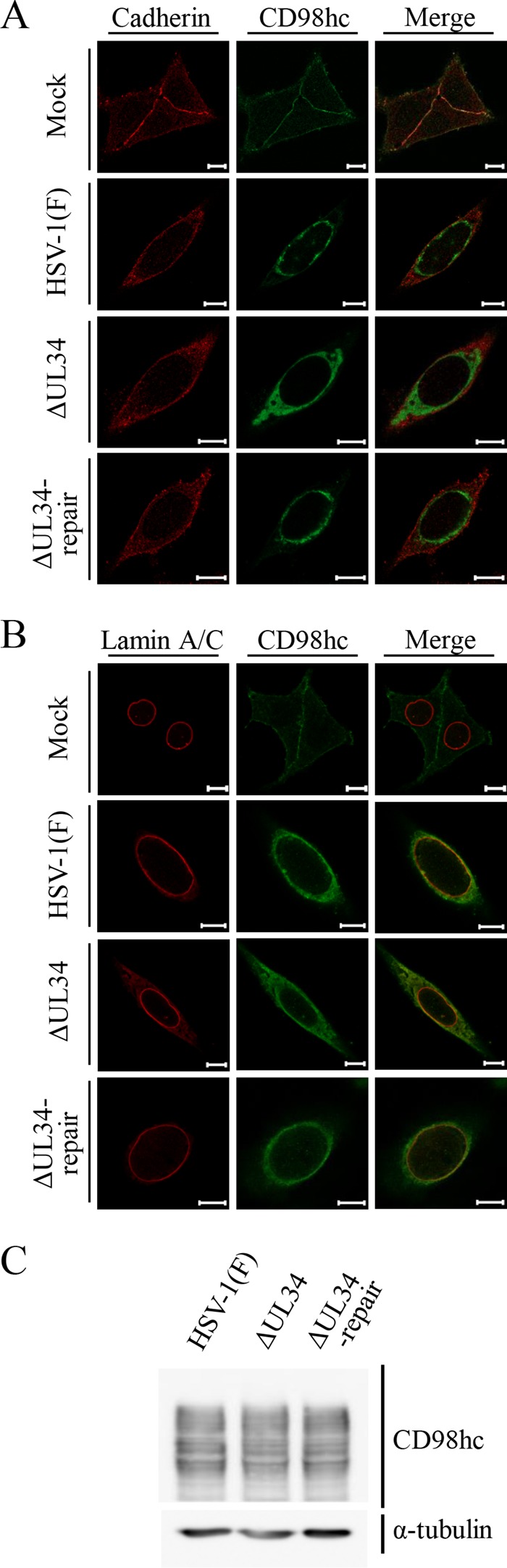
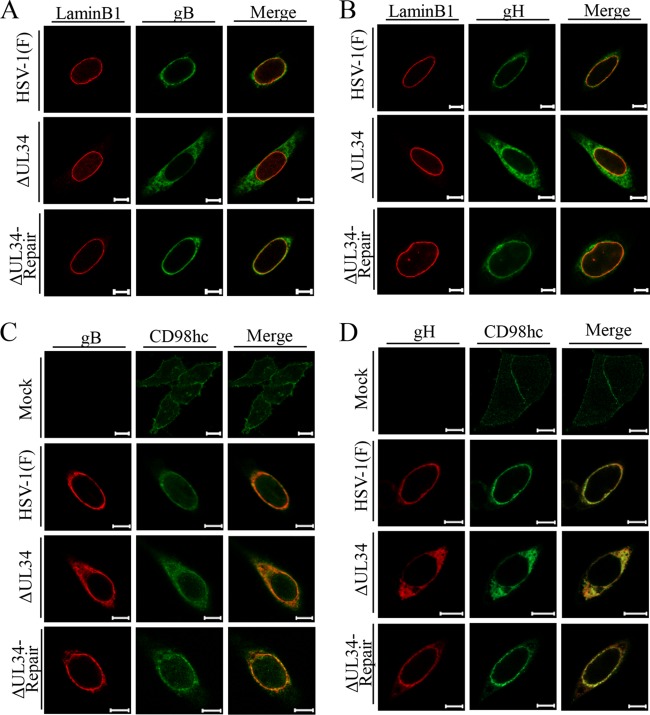
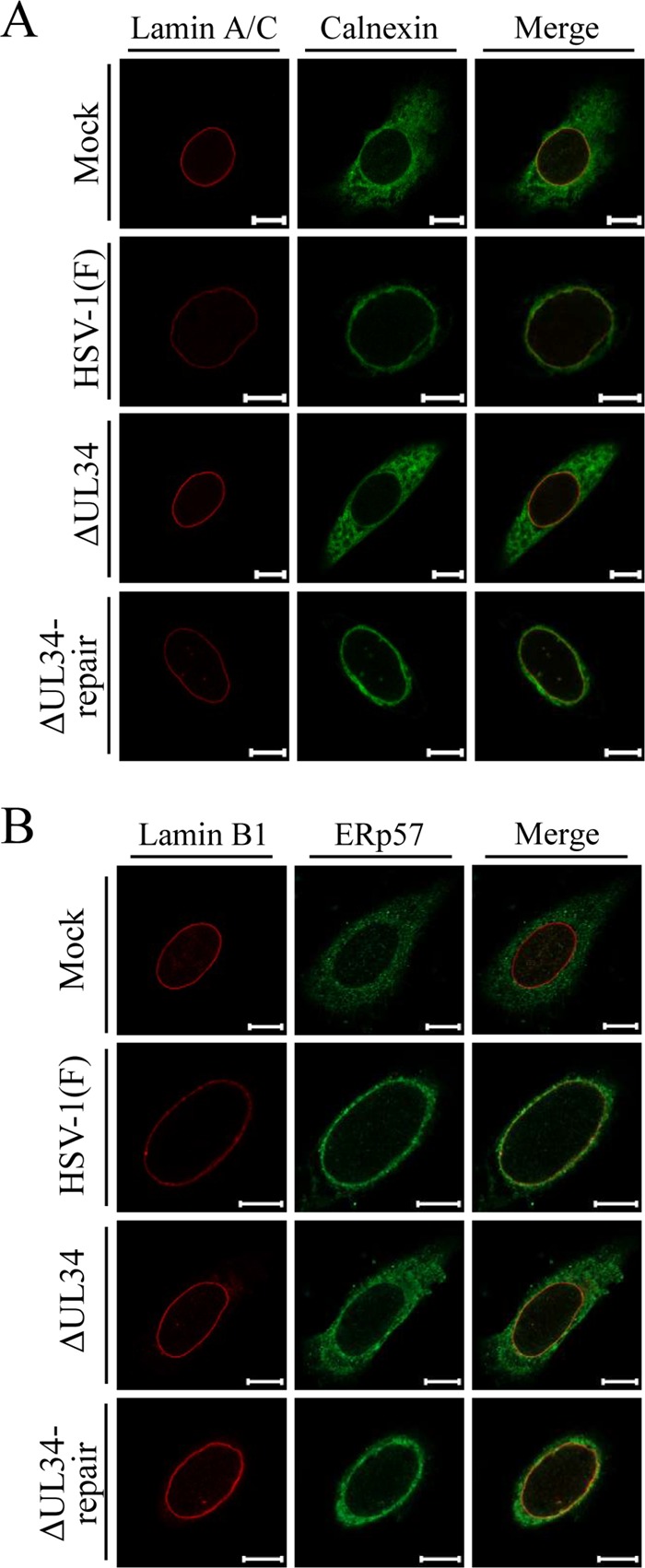
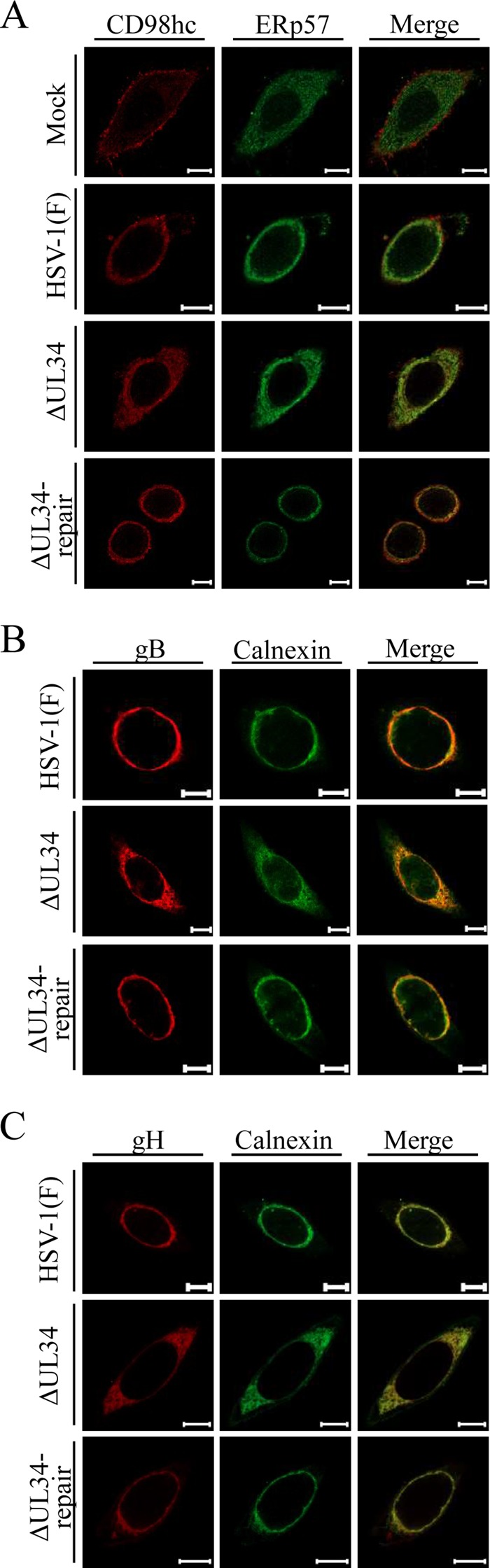
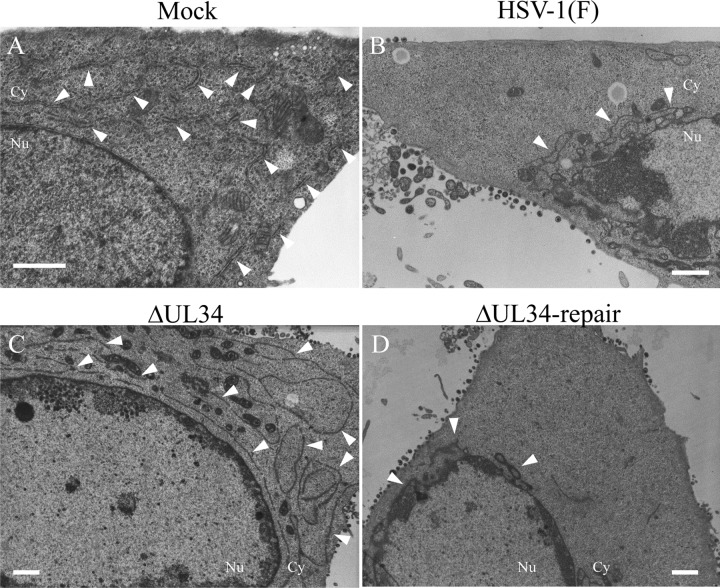
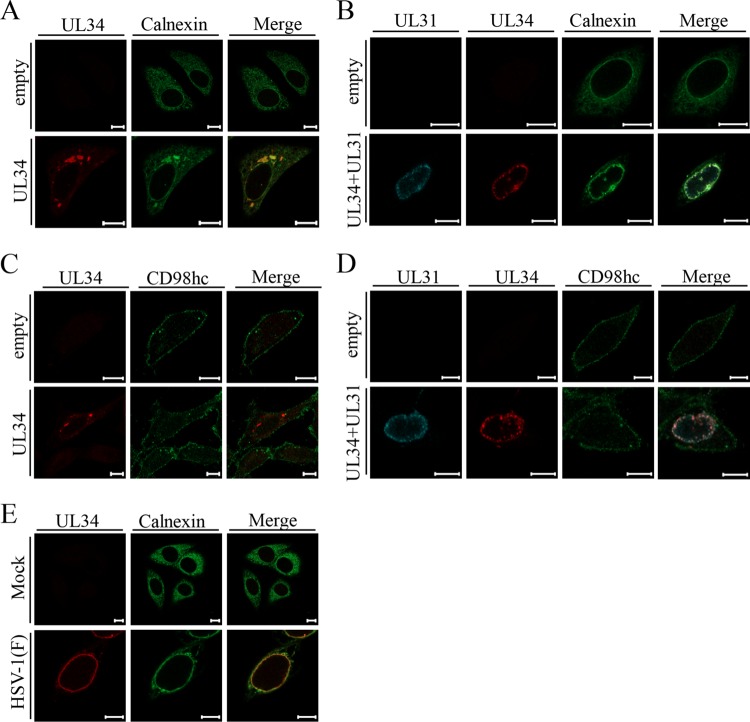
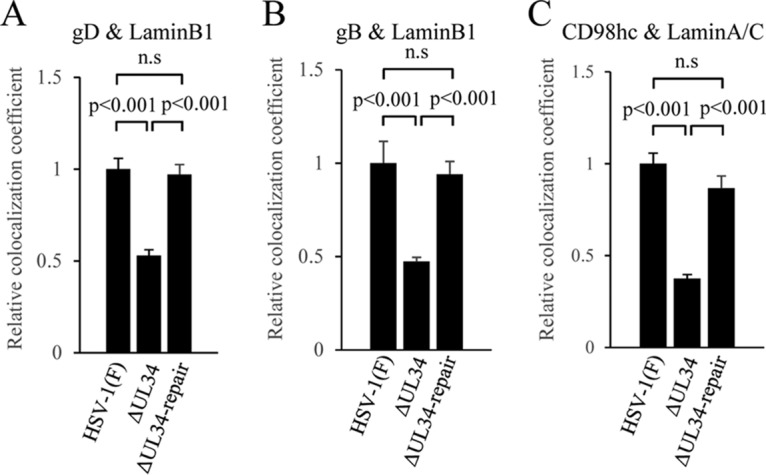
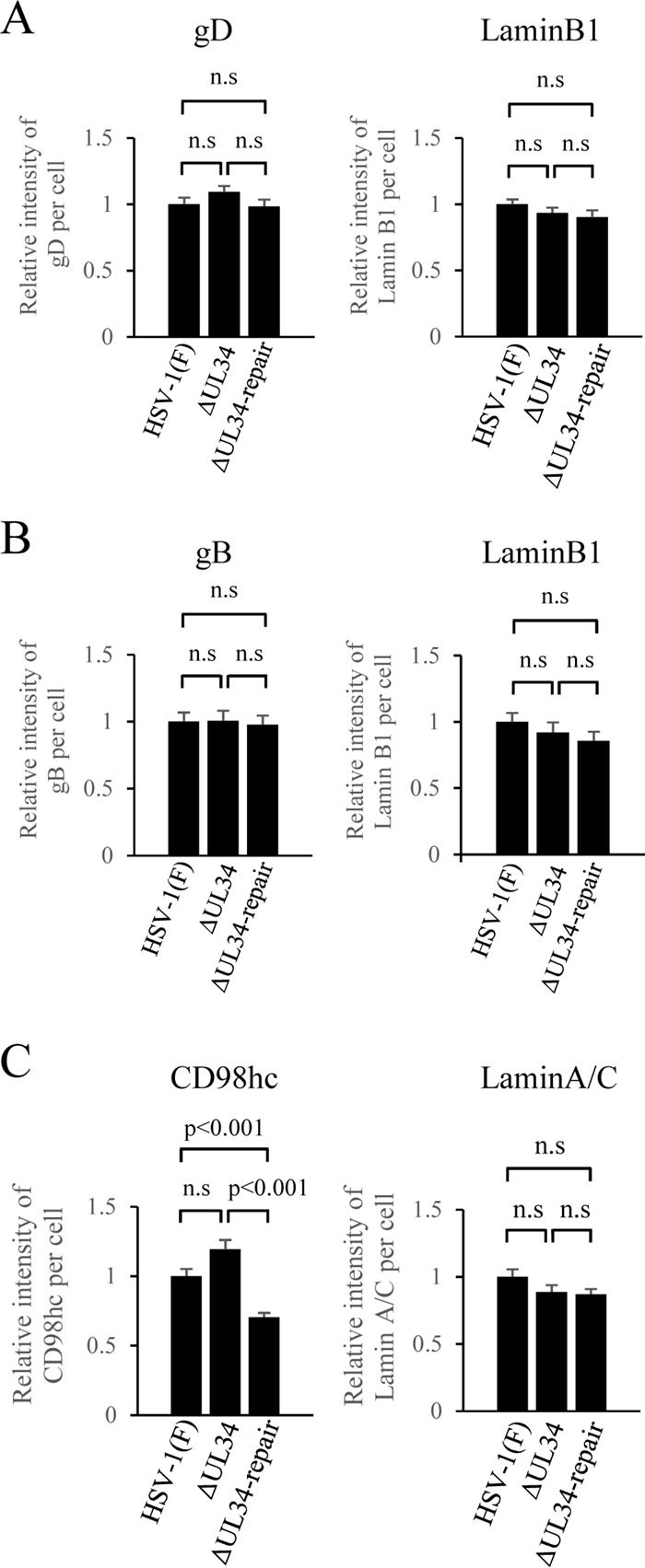
Upon herpes simplex virus 1 (HSV-1) infection, the CD98 heavy chain (CD98hc) is redistributed around the nuclear membrane (NM), where it promotes viral de-envelopment during the nuclear egress of nucleocapsids. In this study, we attempted to identify the factor(s) involved in CD98hc accumulation and demonstrated the following: (i) the null mutation of HSV-1 UL34 caused specific dispersion throughout the cytoplasm of CD98hc and the HSV-1 de-envelopment regulators, glycoproteins B and H (gB and gH); (ii) as observed with CD98hc, gB, and gH, wild-type HSV-1 infection caused redistribution of the endoplasmic reticulum (ER) markers calnexin and ERp57 around the NM, whereas the UL34-null mutation caused cytoplasmic dispersion of these markers; (iii) the ER markers colocalized efficiently with CD98hc, gB, and gH in the presence and absence of UL34 in HSV-1-infected cells; (iv) at the ultrastructural level, wild-type HSV-1 infection caused ER compression around the NM, whereas the UL34-null mutation caused cytoplasmic dispersion of the ER; and (v) the UL34-null mutation significantly decreased the colocalization efficiency of lamin protein markers of the NM with CD98hc and gB. Collectively, these results indicate that HSV-1 infection causes redistribution of the ER around the NM, with resulting accumulation of ER-associated CD98hc, gB, and gH around the NM and that UL34 is required for ER redistribution, as well as for efficient recruitment to the NM of the ER-associated de-envelopment factors. Our study suggests that HSV-1 induces remodeling of the global ER architecture for recruitment of regulators mediating viral nuclear egress to the NM.IMPORTANCE The ER is an important cellular organelle that exists as a complex network extending throughout the cytoplasm. Although viruses often remodel the ER to facilitate viral replication, information on the effects of herpesvirus infections on ER morphological integrity is limited. Here, we showed that HSV-1 infection led to compression of the global ER architecture around the NM, resulting in accumulation of ER-associated regulators associated with nuclear egress of HSV-1 nucleocapsids. We also identified HSV-1 UL34 as a viral factor that mediated ER remodeling. Furthermore, we demonstrated that UL34 was required for efficient targeting of these regulators to the NM. To our knowledge, this is the first report showing that a herpesvirus remodels ER global architecture. Our study also provides insight into the mechanism by which the regulators for HSV-1 nuclear egress are recruited to the NM, where this viral event occurs.
Keywords: ER; herpes simplex virus; organelle structure.
Copyright © 2017 American Society for Microbiology.
Figures
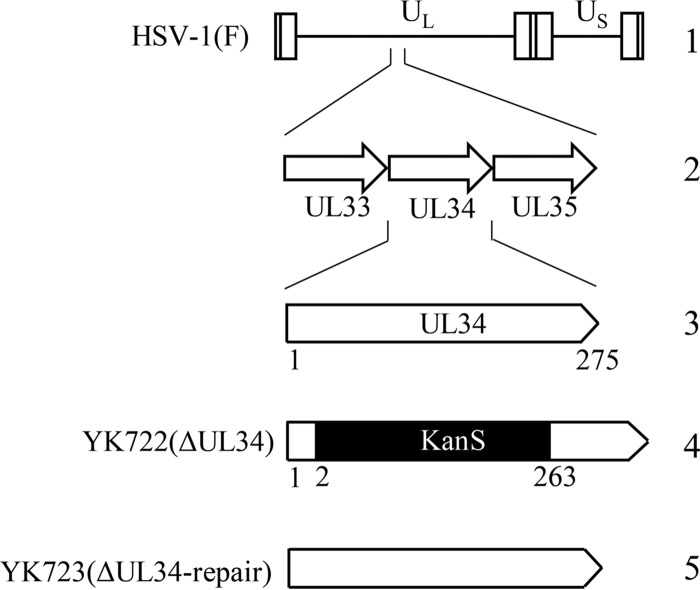

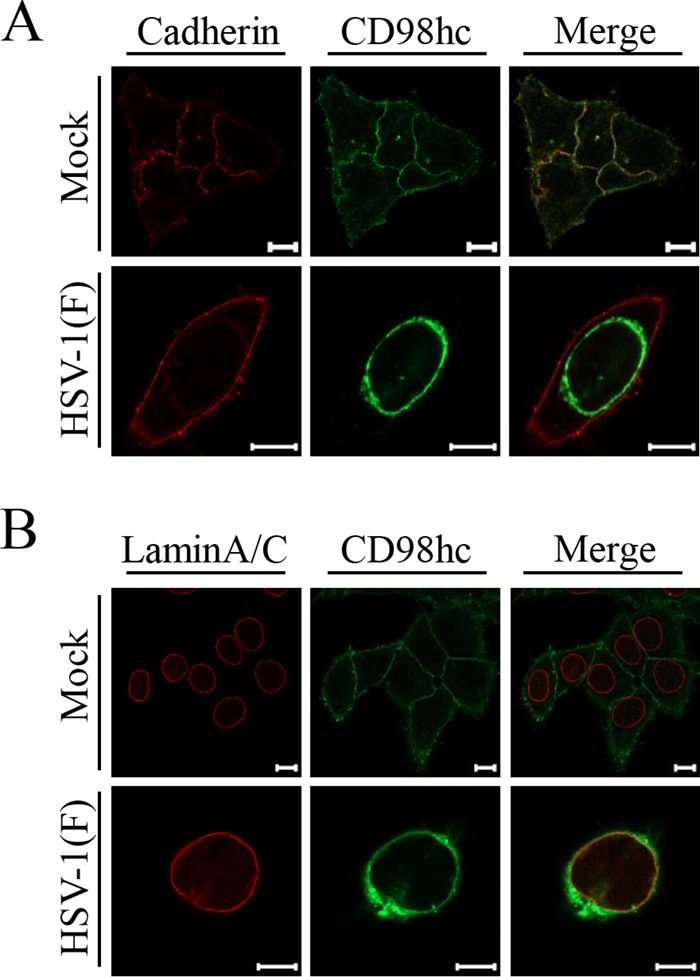
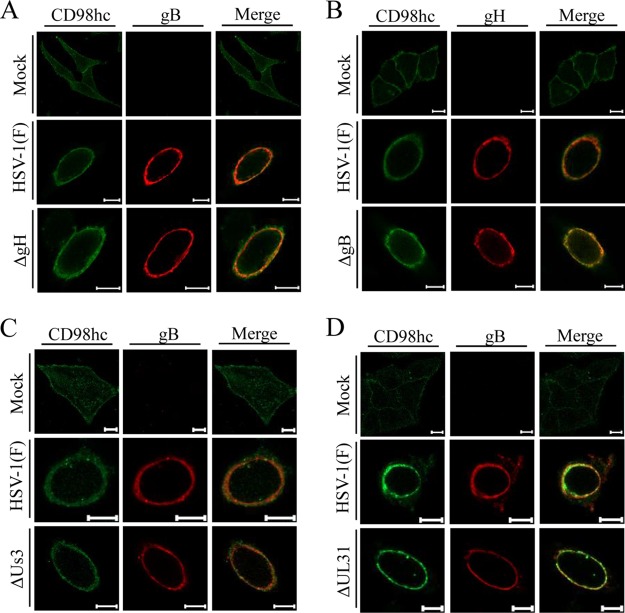
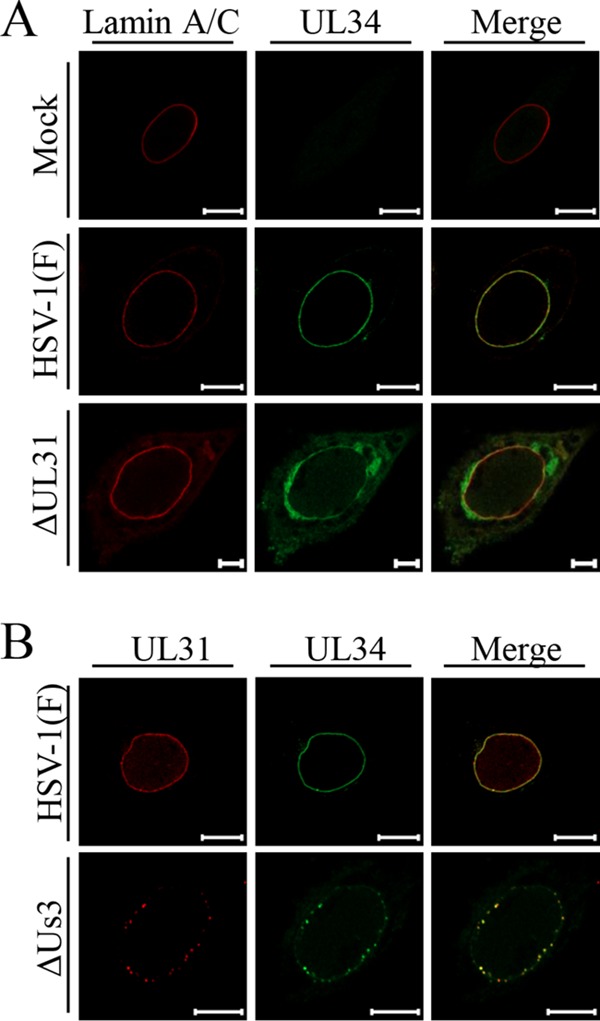
References
-
- Roizman B, Knipe DM, Whitley RJ. 2013. Herpes simplex viruses, p 1823–1897. In Knipe DM, Howley PM, Cohen JI, Griffin DE, Lamb RA, Martin MA, Racaniello VR, Roizman B (ed), Fields virology, 6th ed Lippincott-Williams &Wilkins, Philadelphia, PA.
-
- Reynolds AE, Ryckman BJ, Baines JD, Zhou Y, Liang L, Roller RJ. 2001. U(L)31 and U(L)34 proteins of herpes simplex virus type 1 form a complex that accumulates at the nuclear rim and is required for envelopment of nucleocapsids. J Virol 75:8803–8817. doi: 10.1128/JVI.75.18.8803-8817.2001. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
