

The mystery of membrane organization: composition, regulation and roles of lipid rafts
- PMID: 28356571
- PMCID: PMC5500228
- DOI: 10.1038/nrm.2017.16
The mystery of membrane organization: composition, regulation and roles of lipid rafts
Abstract
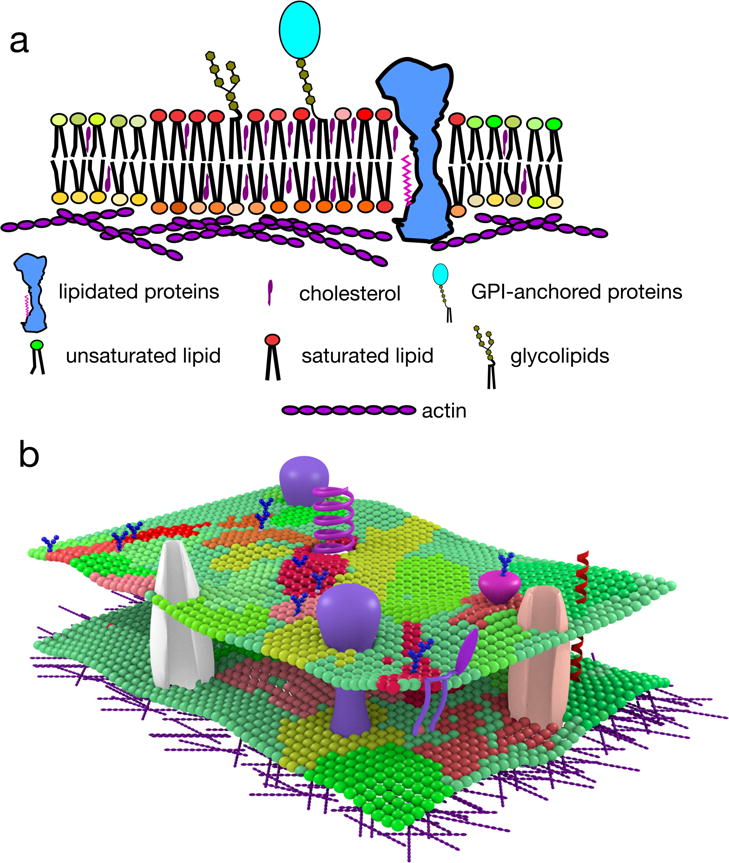
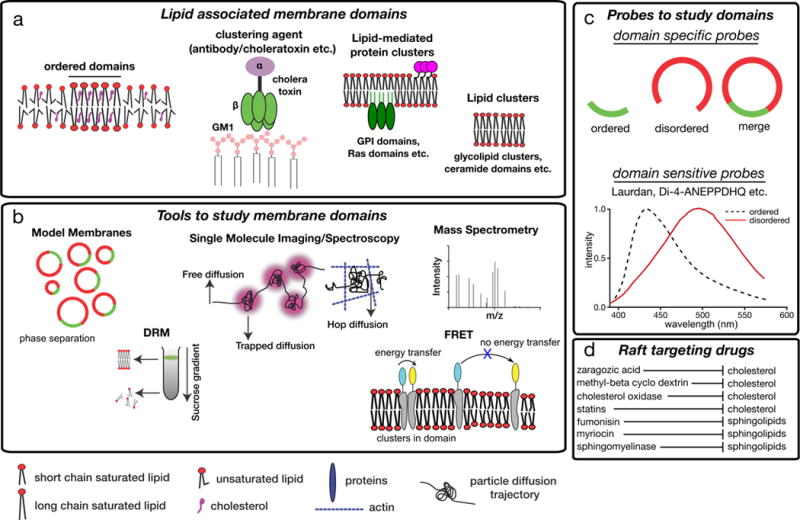
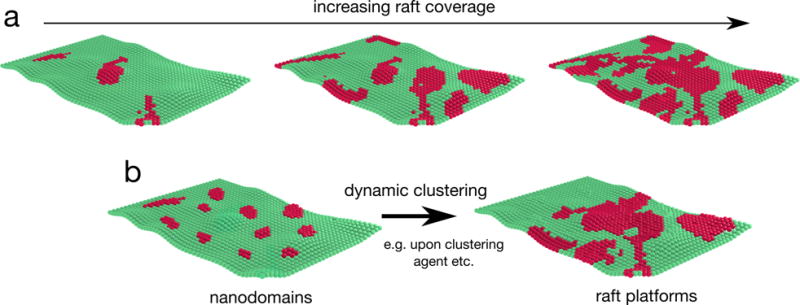
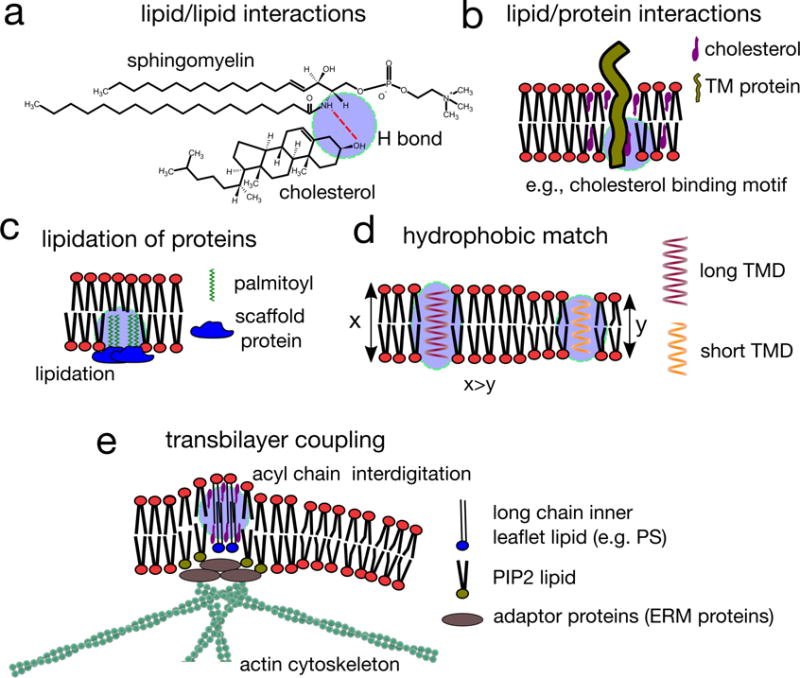
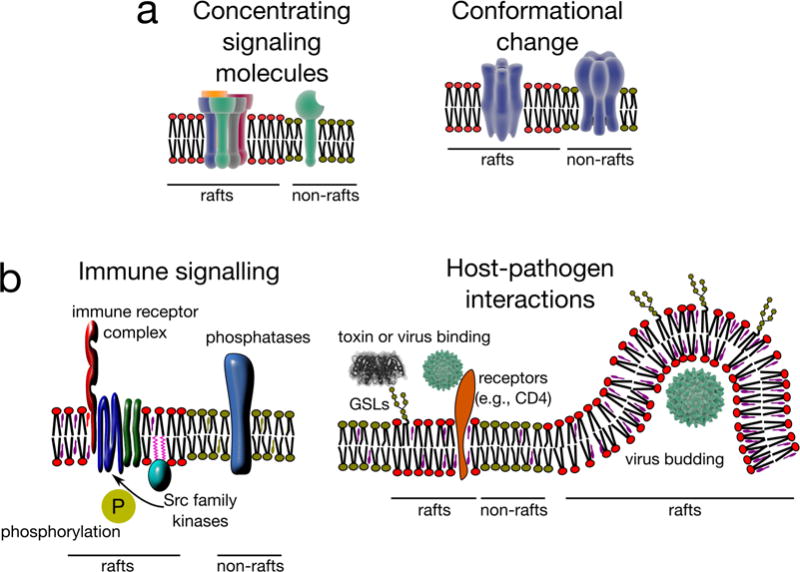
Cellular plasma membranes are laterally heterogeneous, featuring a variety of distinct subcompartments that differ in their biophysical properties and composition. A large number of studies have focused on understanding the basis for this heterogeneity and its physiological relevance. The membrane raft hypothesis formalized a physicochemical principle for a subtype of such lateral membrane heterogeneity, in which the preferential associations between cholesterol and saturated lipids drive the formation of relatively packed (or ordered) membrane domains that selectively recruit certain lipids and proteins. Recent studies have yielded new insights into this mechanism and its relevance in vivo, owing primarily to the development of improved biochemical and biophysical technologies.
Figures
References
-
- Singer SJ, Nicolson GL. The fluid mosaic model of the structure of cell membranes. Science. 1972;175:720–&. - PubMed
-
- Yu J, Fischman DA, Steck TL. Selective solubilization of proteins and phospholipids from red blood cell membranes by nonionic detergents. Journal of Supramolecular Structure. 1973;1:233–248. - PubMed
-
- Ahmed SN, Brown DA, London E. On the origin of sphingolipid/cholesterol-rich detergent-insoluble cell membranes: Physiological concentrations of cholesterol and sphingolipid induce formation of a detergent-insoluble, liquid-ordered lipid phase in model membranes. Biochemistry. 1997;36:10944–10953. - PubMed
-
- Brown DA, Rose JK. Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell. 1992;68:533–544. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
