

Predatory interactions between prey affect patch selection by predators
- PMID: 28356611
- PMCID: PMC5352743
- DOI: 10.1007/s00265-017-2288-2
Predatory interactions between prey affect patch selection by predators
Abstract
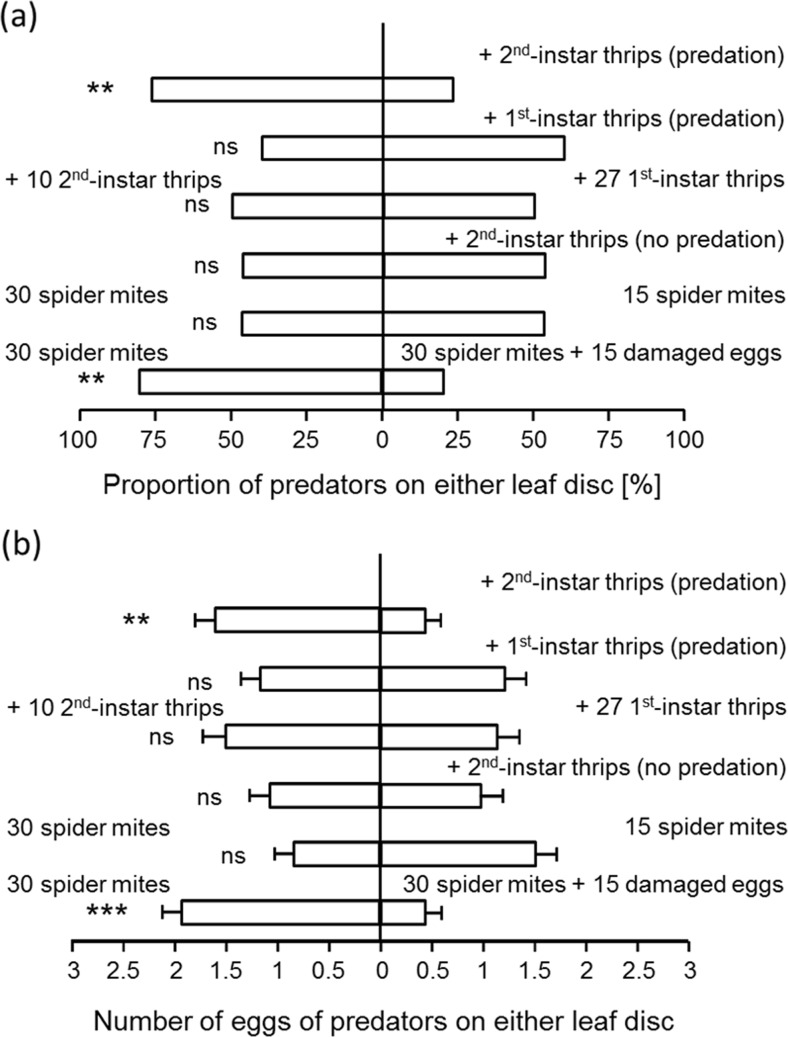
Abstract: When predators can use several prey species as food sources, they are known to select prey according to foraging efficiency and food quality. However, interactions between the prey species may also affect prey choice, and this has received limited attention. The effect of one such interaction, intraguild predation between prey, on patch selection by predators was studied here. The predatory mite Neoseiulus californicus preys on young larvae of the western flower thrips Frankliniella occidentalis and on all stages of the two-spotted spider mite Tetranychus urticae. The two prey species co-occur on several plant species, on which they compete for resources, and western flower thrips feed on eggs of the spider mites. A further complicating factor is that the thrips can also feed on the eggs of the predator. We found that performance of the predatory mite was highest on patches with spider mites, intermediate on patches with spider mites plus thrips larvae and lowest on patches with thrips larvae alone. Patch selection and oviposition preference of predators matched performance: predators preferred patches with spider mites over patches with spider mites plus thrips. Patches with thrips only were not significantly more attractive than empty patches. We also investigated the cues involved in patch selection and found that the attractiveness of patches with spider mites was significantly reduced by the presence of cues associated with killed spider mite eggs. This explains the reduced attractiveness of patches with both prey. Our results point at the importance of predatory interactions among prey species for patch selection by predators.
Significance statement: Patch selection by predators is known to be affected by factors such as prey quality, the presence of competitors and predators, but little is known on the effects of interactions among prey species present on patch selection. In this paper, we show that patch selection by a predator is affected by such interactions, specifically by the feeding of one prey species on eggs of the other.
Keywords: Behaviour; Competition; Neoseiulus californicus; Prey-prey interactions; Shared predator; Spider mites; Thrips.
Figures



Similar articles
-
Intraguild interactions among three spider mite predators: predation preference and effects on juvenile development and oviposition.Exp Appl Acarol. 2015 Dec;67(4):493-505. doi: 10.1007/s10493-015-9974-4. Exp Appl Acarol. 2015. PMID: 26462926
-
Predatory mites avoid ovipositing near counterattacking prey.Exp Appl Acarol. 2001;25(8):613-23. doi: 10.1023/a:1016100212909. Exp Appl Acarol. 2001. PMID: 12171271
-
Intraguild Prey Served as Alternative Prey for Intraguild Predators in a Reciprocal Predator Guild between Neoseiulus barkeri and Scolothrips takahashii.Insects. 2023 Jun 16;14(6):561. doi: 10.3390/insects14060561. Insects. 2023. PMID: 37367377 Free PMC article.
-
Residual effect of commonly used fungicides in strawberries on Amblyseius swirskii, Neoseiulus cucumeris, and Neoseiulus californicus (Mesostigmata: Phytoseiidae).Exp Appl Acarol. 2024 Aug;93(2):253-272. doi: 10.1007/s10493-024-00928-1. Epub 2024 Jun 13. Exp Appl Acarol. 2024. PMID: 38869730 Free PMC article. Review.
-
Thysanoptera as predators: their diversity and significance as biological control agents.Pest Manag Sci. 2022 Dec;78(12):5057-5070. doi: 10.1002/ps.7176. Epub 2022 Sep 26. Pest Manag Sci. 2022. PMID: 36087293 Review.
Cited by
-
Leaf trichome-mediated predator effects on the distribution of herbivorous mites within a kidney bean plant.Exp Appl Acarol. 2024 Jun;93(1):155-167. doi: 10.1007/s10493-024-00915-6. Epub 2024 Apr 11. Exp Appl Acarol. 2024. PMID: 38600348
-
Sister predatory mites collectively protect their eggs against predators.Oecologia. 2024 Mar;204(3):653-660. doi: 10.1007/s00442-024-05521-2. Epub 2024 Mar 9. Oecologia. 2024. PMID: 38461225
-
Role of kin recognition in oviposition preference and cannibalism by the predatory mite Gynaeseius liturivorus.Exp Appl Acarol. 2018 Oct;76(2):149-160. doi: 10.1007/s10493-018-0298-z. Epub 2018 Sep 24. Exp Appl Acarol. 2018. PMID: 30251068
References
-
- Abrams PA. Implications of dramatically variable traits for identifying, classifying and measuring direct and indirect effects in ecological communities. Am Nat. 1995;146:112–134. doi: 10.1086/285789. - DOI
-
- Adler FR, Richards SA, de Roos AM. Patterns of patch rejection in size-structured populations: beyond the ideal free distribution and size segregation. Evol Ecol Res. 2001;3:805–827.
-
- Agrawal AA. Specificity of induced resistance in wild radish: causes and consequences for two specialist and two generalist caterpillars. Oikos. 2000;89:493–500. doi: 10.1034/j.1600-0706.2000.890308.x. - DOI
-
- Agrawal AA, Klein CN. What omnivores eat: direct effects of induced plant resistance on herbivores and indirect consequences for diet selection by omnivores. J Anim Ecol. 2000;69:525–535. doi: 10.1046/j.1365-2656.2000.00416.x. - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources