

Time resolved DNA occupancy dynamics during the respiratory oscillation uncover a global reset point in the yeast growth program
- PMID: 28357254
- PMCID: PMC5349131
- DOI: 10.15698/mic2014.09.166
Time resolved DNA occupancy dynamics during the respiratory oscillation uncover a global reset point in the yeast growth program
Abstract
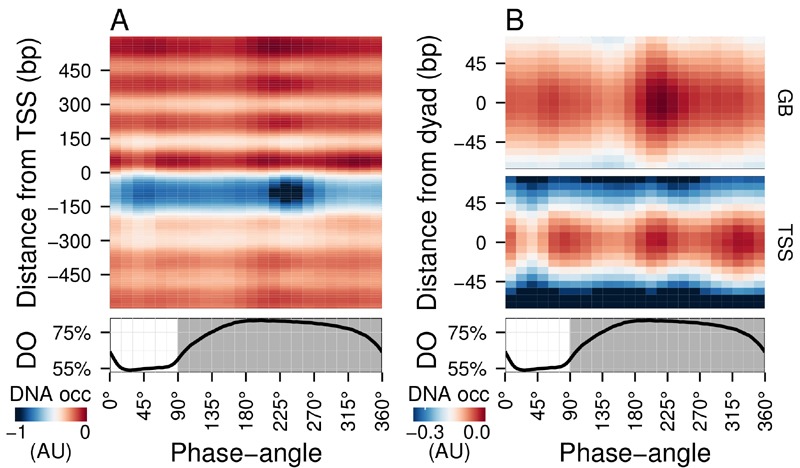
The structural dynamics of chromatin have been implicated in the regulation of fundamental eukaryotic processes, such as DNA transcription, replication and repair. Although previous studies have revealed that the chromatin landscape, nucleosome remodeling and histone modification events are intimately tied into cellular energetics and redox state, few studies undertake defined time-resolved measurements of these state variables. Here, we use metabolically synchronous, continuously-grown yeast cultures to measure DNA occupancy and track global patterns with respect to the metabolic state of the culture. Combined with transcriptome analyses and ChIP-qPCR experiments, these paint an intriguing picture where genome-wide nucleosome focusing occurs during the recovery of energy charge, followed by clearance of the promoter regions and global transcriptional slow-down, thus indicating a nucleosome-mediated "reset point" for the cycle. The reset begins at the end of the catabolic and stress-response transcriptional programs and ends prior to the start of the anabolic and cell-growth transcriptional program, and the histones on genes from both the catabolic and anabolic superclusters are deacetylated.
Keywords: anabolism; catabolism; chromatin dynamics; energetics; histone modification; respiratory oscillation; transcription regulation.
Conflict of interest statement
Conflict of interest: The authors declare no conflict of interest.
Figures




References
-
- Holde KE. New York: Springer; 1989. Chromatin. - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases