

A Novel Staphylococcus Podophage Encodes a Unique Lysin with Unusual Modular Design
- PMID: 28357414
- PMCID: PMC5362749
- DOI: 10.1128/mSphere.00040-17
A Novel Staphylococcus Podophage Encodes a Unique Lysin with Unusual Modular Design
Abstract
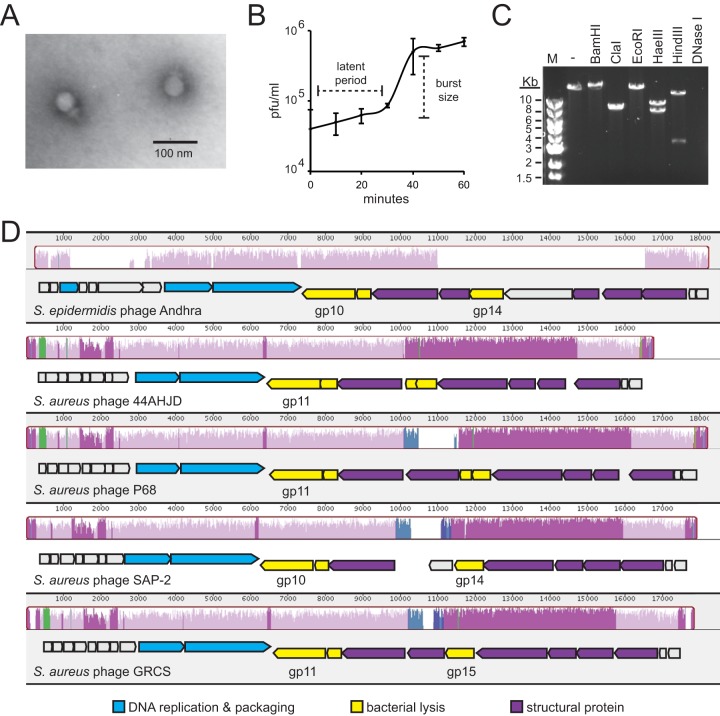
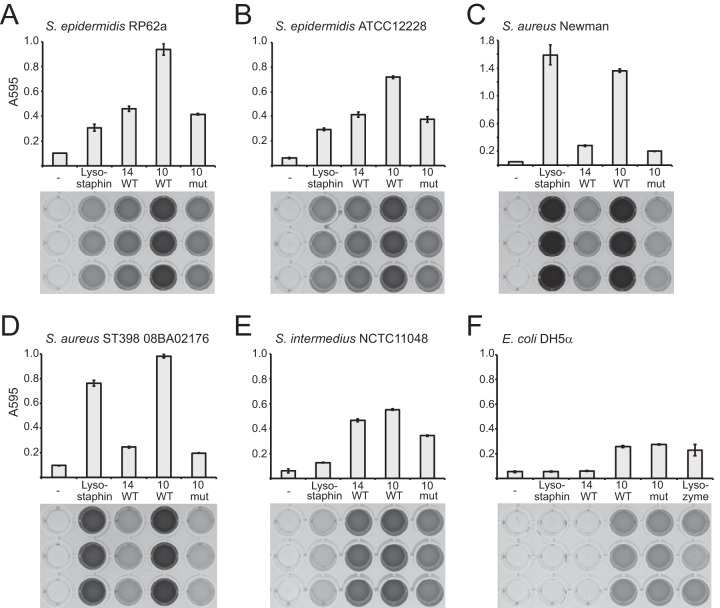
Drug-resistant staphylococci, particularly Staphylococcus aureus and Staphylococcus epidermidis, are leading causes of hospital-acquired infections. Bacteriophages and their peptidoglycan hydrolytic enzymes (lysins) are currently being explored as alternatives to conventional antibiotics; however, only a limited diversity of staphylococcal phages and their lysins has yet been characterized. Here, we describe a novel staphylococcal phage and its lysins. Bacteriophage Andhra is the first reported S. epidermidis phage belonging to the family Podoviridae. Andhra possesses an 18,546-nucleotide genome with 20 open reading frames. BLASTp searches revealed that gene product 10 (gp10) and gp14 harbor putative catalytic domains with predicted peptidase and amidase activities, characteristic functions of phage lysins. We purified these proteins and show that both Andhra_gp10 and Andhra_gp14 inhibit growth and degrade cell walls of diverse staphylococci, with Andhra_gp10 exhibiting more robust activity against the panel of cell wall substrates tested. Site-directed mutagenesis of its predicted catalytic residues abrogated the activity of Andhra_gp10, consistent with the presence of a catalytic CHAP domain on its C terminus. The active site location combined with the absence of an SH3b cell wall binding domain distinguishes Andhra_gp10 from the majority of staphylococcal lysins characterized to date. Importantly, close homologs of Andhra_gp10 are present in related staphylococcal podophages, and we propose that these constitute a new class of phage-encoded lysins. Altogether, our results reveal insights into the biology of a rare family of staphylococcal phages while adding to the arsenal of antimicrobials with potential for therapeutic use. IMPORTANCE The spread of antibiotic resistance among bacterial pathogens is inciting a global public health crisis. Drug-resistant Staphylococcus species, especially S. aureus and S. epidermidis, have emerged in both hospital and community settings, underscoring the urgent need for new strategies to combat staphylococcal infections. Bacterial viruses (phages) and the enzymes that they use to degrade bacterial cell walls (lysins) show promise as alternative antimicrobials; however, only a limited variety of staphylococcal phages and their lysins have yet been identified. Here, we report the discovery and characterization of a novel staphylococcal phage, Andhra. We show that Andhra encodes two lysins (Andhra_gp10 and Andhra_gp14) that inhibit growth and degrade the cell walls of diverse staphylococci, including S. aureus and S. epidermidis strains. Andhra and its unique lysins add to the arsenal of antimicrobials with potential for therapeutic use.
Keywords: Staphylococcus; antimicrobial agents; bacteriophage lysis; bacteriophage therapy; bacteriophages.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
