

Functional inhibition of acid sphingomyelinase by Fluphenazine triggers hypoxia-specific tumor cell death
- PMID: 28358364
- PMCID: PMC5386533
- DOI: 10.1038/cddis.2017.130
Functional inhibition of acid sphingomyelinase by Fluphenazine triggers hypoxia-specific tumor cell death
Abstract
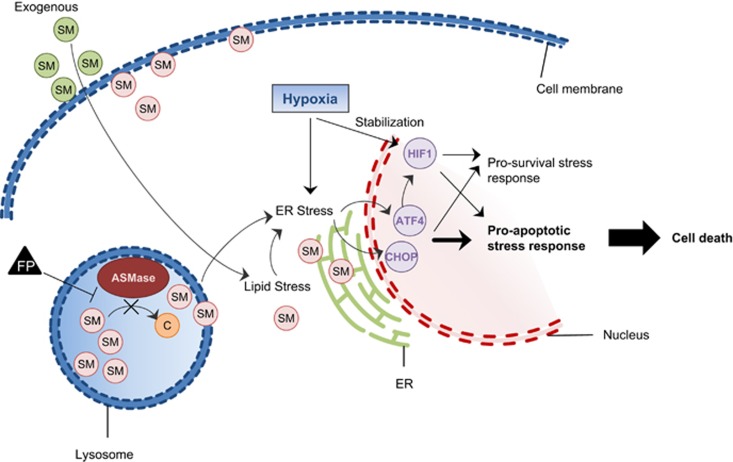
Owing to lagging or insufficient neo-angiogenesis, hypoxia is a feature of most solid tumors. Hypoxic tumor regions contribute to resistance against antiproliferative chemotherapeutics, radiotherapy and immunotherapy. Targeting cells in hypoxic tumor areas is therefore an important strategy for cancer treatment. Most approaches for targeting hypoxic cells focus on the inhibition of hypoxia adaption pathways but only a limited number of compounds with the potential to specifically target hypoxic tumor regions have been identified. By using tumor spheroids in hypoxic conditions as screening system, we identified a set of compounds, including the phenothiazine antipsychotic Fluphenazine, as hits with novel mode of action. Fluphenazine functionally inhibits acid sphingomyelinase and causes cellular sphingomyelin accumulation, which induces cancer cell death specifically in hypoxic tumor spheroids. Moreover, we found that functional inhibition of acid sphingomyelinase leads to overactivation of hypoxia stress-response pathways and that hypoxia-specific cell death is mediated by the stress-responsive transcription factor ATF4. Taken together, the here presented data suggest a novel, yet unexplored mechanism in which induction of sphingolipid stress leads to the overactivation of hypoxia stress-response pathways and thereby promotes their pro-apoptotic tumor-suppressor functions to specifically kill cells in hypoxic tumor areas.
Conflict of interest statement
All authors are employees of Bayer AG.
Figures






References
-
- Kyle AH, Baker JH, Minchinton AI. Targeting quiescent tumor cells via oxygen and IGF-I supplementation. Cancer Res 2012; 72: 801–809. - PubMed
-
- Semenza GL. Oxygen sensing, hypoxia-inducible factors, and disease pathophysiology. Annu Rev Pathol 2014; 9: 47–71. - PubMed
-
- Koumenis C, Wouters BG. "Translating" tumor hypoxia: unfolded protein response (UPR)-dependent and UPR-independent pathways. Mol Cancer Res 2006; 4: 423–436. - PubMed
-
- Vaupel P, Mayer A. Hypoxia in cancer: significance and impact on clinical outcome. Cancer Metastasis Rev 2007; 26: 225–239. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
