

Phenotypically silent Cre recombination within the postnatal ventricular conduction system
- PMID: 28358866
- PMCID: PMC5373586
- DOI: 10.1371/journal.pone.0174517
Phenotypically silent Cre recombination within the postnatal ventricular conduction system
Abstract
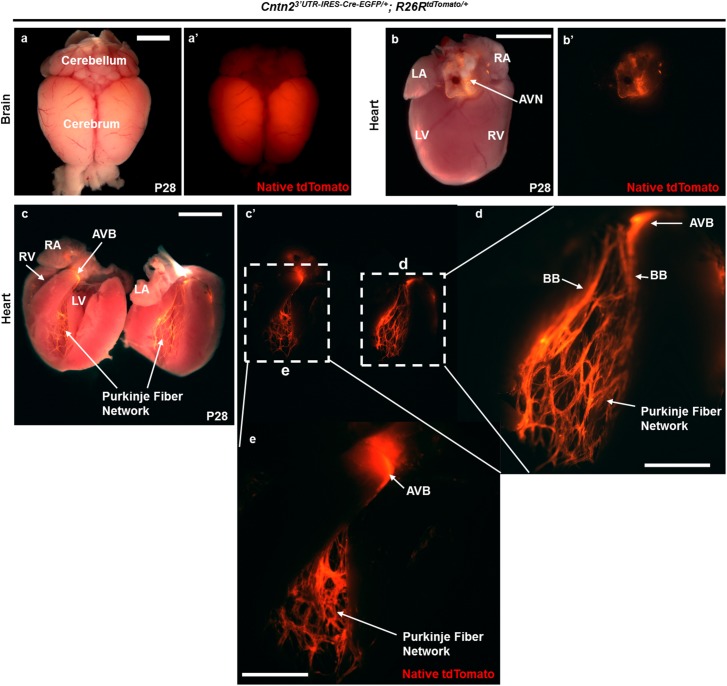
The cardiac conduction system (CCS) is composed of specialized cardiomyocytes that initiate and maintain cardiac rhythm. Any perturbation to the normal sequence of electrical events within the heart can result in cardiac arrhythmias. To understand how cardiac rhythm is established at the molecular level, several genetically modified mouse lines expressing Cre recombinase within specific CCS compartments have been created. In general, Cre driver lines have been generated either by homologous recombination of Cre into an endogenous locus or Cre expression driven by a randomly inserted transgene. However, haploinsufficiency of the endogenous gene compromises the former approach, while position effects negatively impact the latter. To address these limitations, we generated a Cre driver line for the ventricular conduction system (VCS) that preserves endogenous gene expression by targeting the Contactin2 (Cntn2) 3' untranslated region (3'UTR). Here we show that Cntn23'UTR-IRES-Cre-EGFP/+ mice recombine floxed alleles within the VCS and that Cre expression faithfully recapitulates the spatial distribution of Cntn2 within the heart. We further demonstrate that Cre expression initiates after birth with preservation of native Cntn2 protein. Finally, we show that Cntn23'UTR-IRES-Cre-EGFP/+ mice maintain normal cardiac mechanical and electrical function. Taken together, our results establish a novel VCS-specific Cre driver line without the adverse consequences of haploinsufficiency or position effects. We expect that our new mouse line will add to the accumulating toolkit of CCS-specific mouse reagents and aid characterization of the cell-autonomous molecular circuitry that drives VCS maintenance and function.
Conflict of interest statement
Figures





References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
