

Breathing control center neurons that promote arousal in mice
- PMID: 28360327
- PMCID: PMC5505554
- DOI: 10.1126/science.aai7984
Breathing control center neurons that promote arousal in mice
Abstract
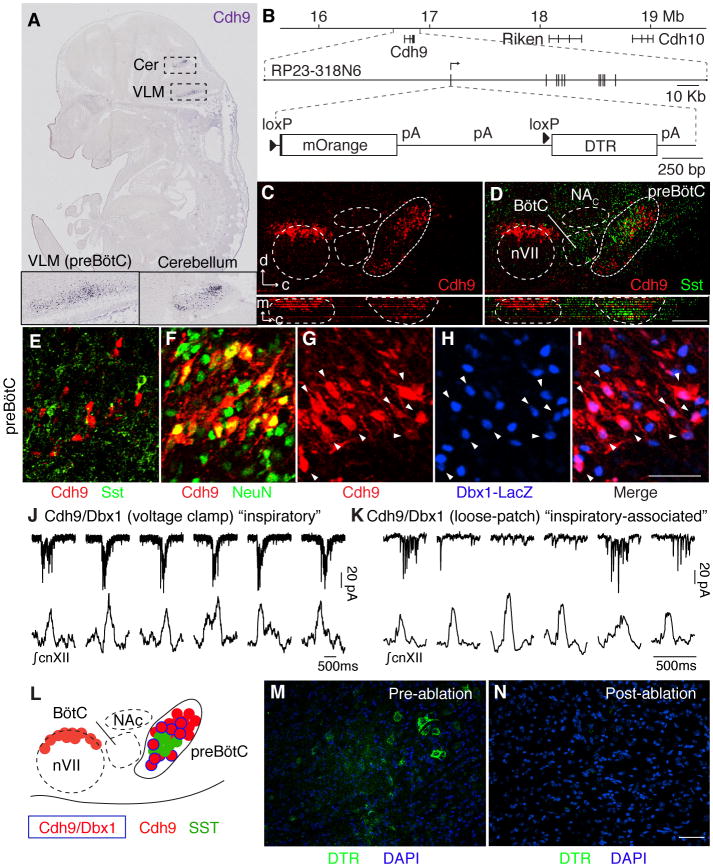
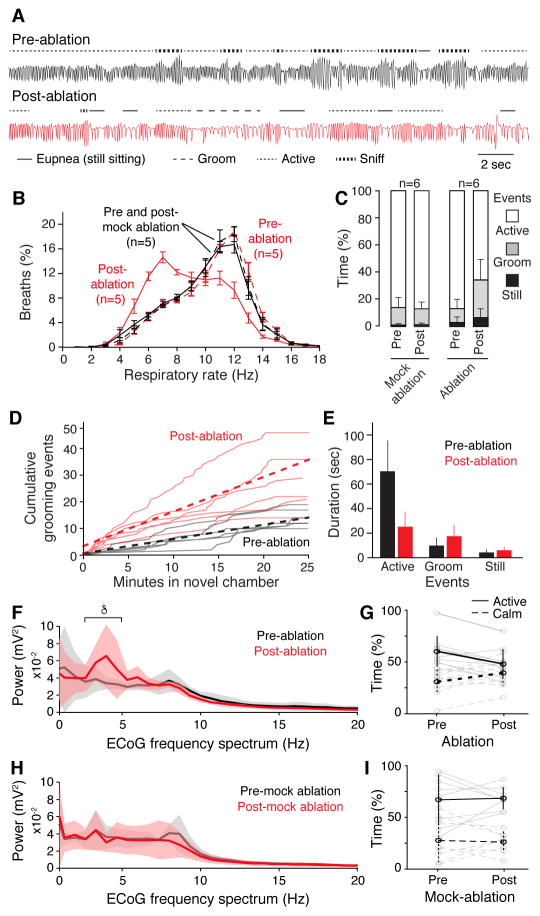
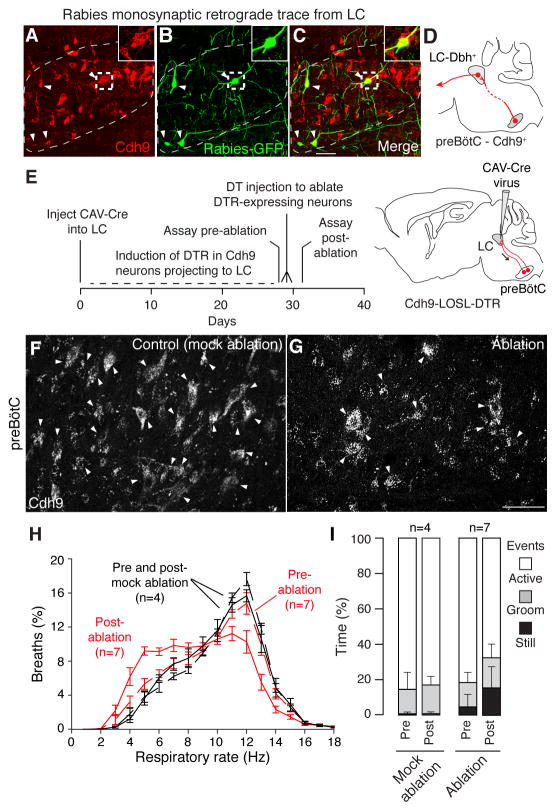
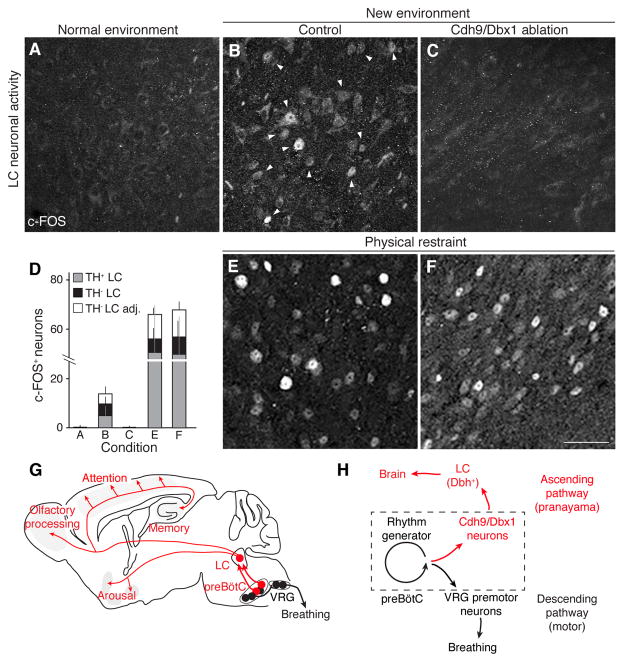
Slow, controlled breathing has been used for centuries to promote mental calming, and it is used clinically to suppress excessive arousal such as panic attacks. However, the physiological and neural basis of the relationship between breathing and higher-order brain activity is unknown. We found a neuronal subpopulation in the mouse preBötzinger complex (preBötC), the primary breathing rhythm generator, which regulates the balance between calm and arousal behaviors. Conditional, bilateral genetic ablation of the ~175 Cdh9/Dbx1 double-positive preBötC neurons in adult mice left breathing intact but increased calm behaviors and decreased time in aroused states. These neurons project to, synapse on, and positively regulate noradrenergic neurons in the locus coeruleus, a brain center implicated in attention, arousal, and panic that projects throughout the brain.
Copyright © 2017, American Association for the Advancement of Science.
Figures
Comment in
-
Breathing to inspire and arouse.Science. 2017 Mar 31;355(6332):1370-1371. doi: 10.1126/science.aan1466. Science. 2017. PMID: 28360280 Free PMC article. No abstract available.
References
-
- Brown RP, Gerbarg PL. Yoga breathing, meditation, and longevity. Ann N Y Acad Sci. 2009;1172:54–62. - PubMed
-
- Brown RP, Gerbarg PL. Sudarshan Kriya Yogic breathing in the treatment of stress, anxiety, and depression: part II-clinical applications and guidelines. J Altern Complement Med. 2005;11:711–717. - PubMed
-
- Nardi E, Freire RC, Zin WA. Panic disorder and control of breathing. Respir Physiol Neurobiol. 2009;167:133–143. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
