

Interplay of DNA methyltransferase 1 and EZH2 through inactivation of Stat3 contributes to β-elemene-inhibited growth of nasopharyngeal carcinoma cells
- PMID: 28360411
- PMCID: PMC5428779
- DOI: 10.1038/s41598-017-00626-6
Interplay of DNA methyltransferase 1 and EZH2 through inactivation of Stat3 contributes to β-elemene-inhibited growth of nasopharyngeal carcinoma cells
Abstract
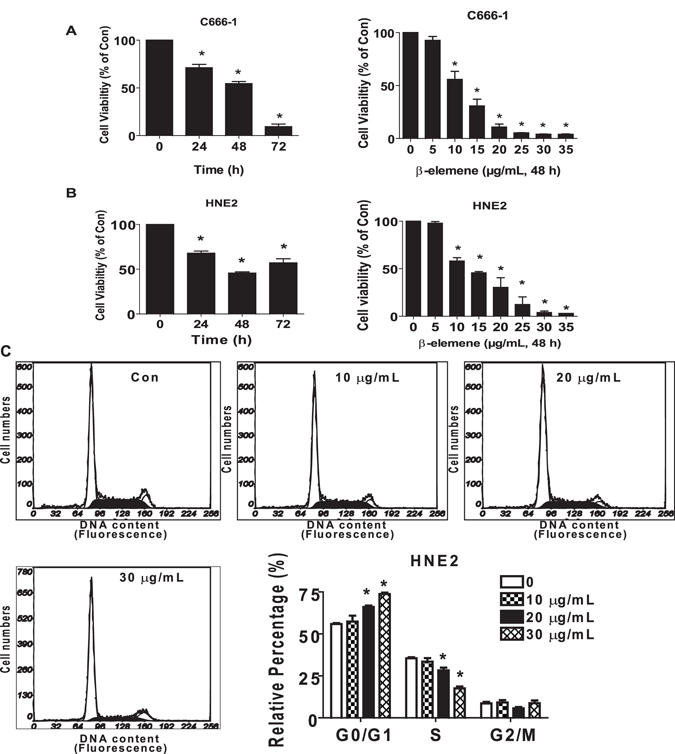
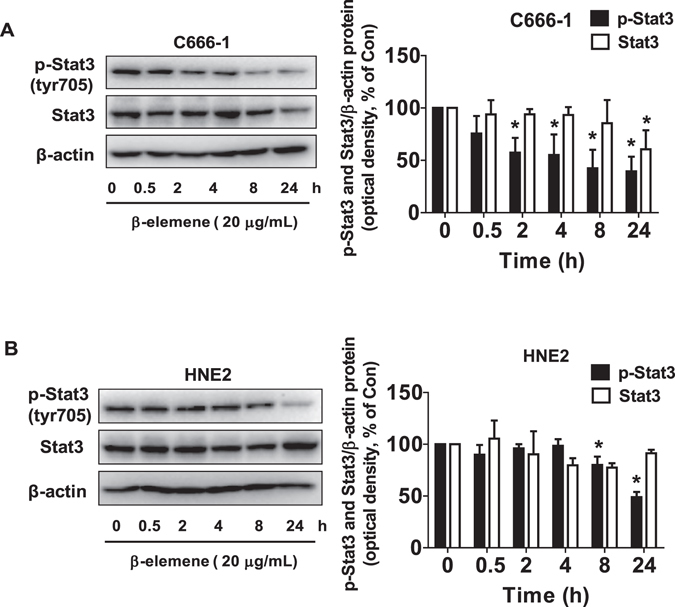
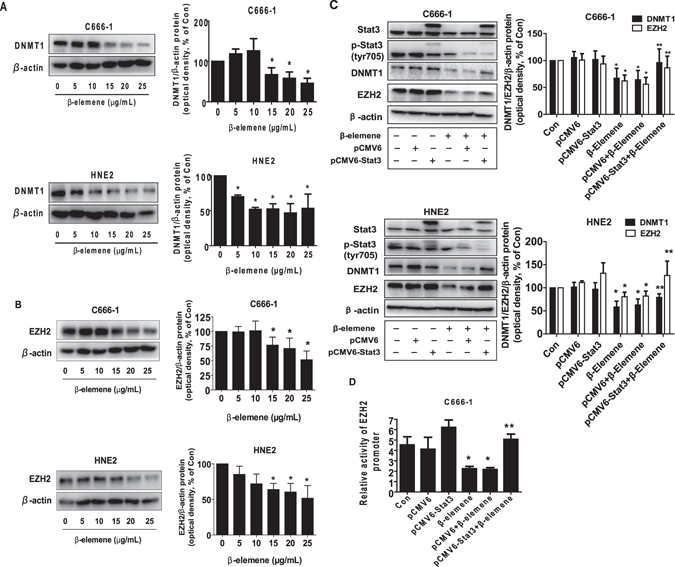
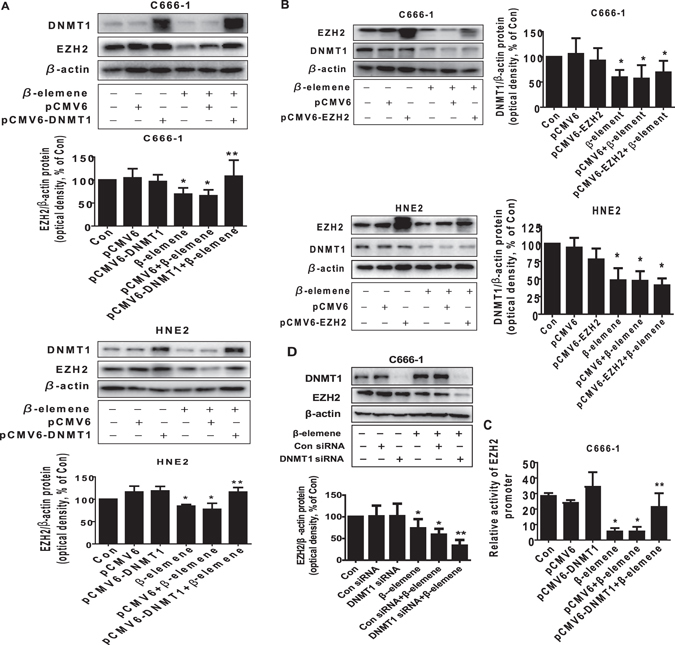
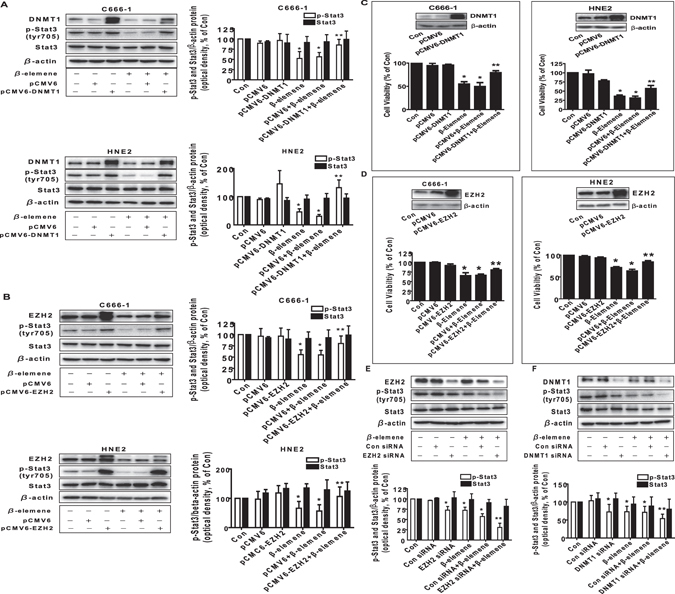
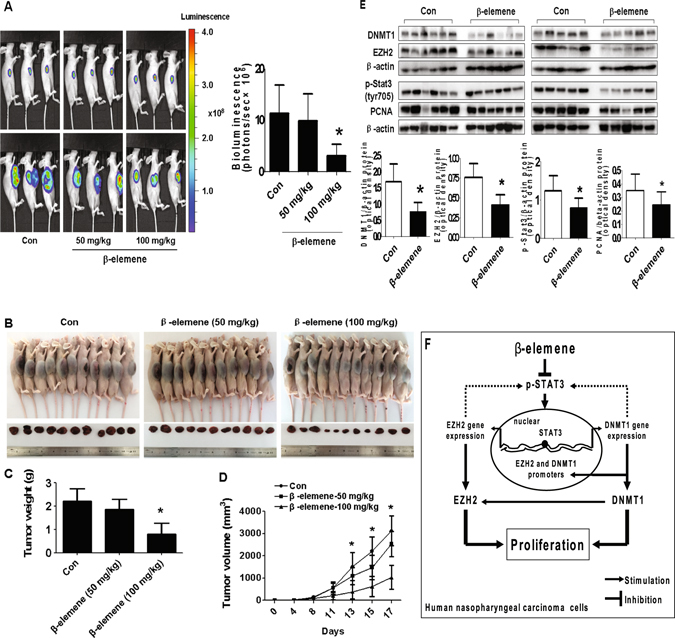
β-elemene, a compound extracted from Curcuma wenyujin plant, exhibits anticancer activity in many cancer types. However, the detailed mechanism by which β-elemene inhibits growth of nasopharyngeal carcinoma (NPC) cells remains unknown. We showed that β-elemene reduced phosphorylation of signal transducer and activator of transcription 3 (Stat3), and protein expressions of DNA methyltransferase 1 (DNMT1) and enhancer of zeste homolog 2 (EZH2). Exogenously expressed Stat3 antagonized the effect of β-elemene on DNMT1 and EZH2 expressions. Furthermore, overexpressions of DNMT1 and EZH2 reversed the effect of β-elemene on phosphorylation of Stat3 and cell growth inhibition. Intriguingly, exogenously expressed DNMT1 overcame β-elemene-inhibited EZH2 protein expression and promoter activity. On the contrary, silencing of EZH2 and DNMT1 genes feedback strengthened the effect of β-elemene on phosphorylation of Stat3. Consistent with this, β-elemene inhibited tumor growth, phosphorylation of Stat3, expressions of DNMT1 and EZH2 in a mouse xenograft model. Collectively, this study shows that β-elemene inhibits NPC cell growth via inactivation of Stat3, and reduces DNMT1 and EZH2 expressions. The interplay of DNMT1 and EZH2, and the mutual regulations among Stat3, EZH2 and DNMT1 contribute to the overall responses of β-elemene. This study uncovers a novel mechanism by which β-elemene inhibits growth of NPC cells.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Inhibition of EZH2 via activation of SAPK/JNK and reduction of p65 and DNMT1 as a novel mechanism in inhibition of human lung cancer cells by polyphyllin I.J Exp Clin Cancer Res. 2016 Jul 16;35(1):112. doi: 10.1186/s13046-016-0388-x. J Exp Clin Cancer Res. 2016. PMID: 27421653 Free PMC article.
-
β-elemene inhibited expression of DNA methyltransferase 1 through activation of ERK1/2 and AMPKα signalling pathways in human lung cancer cells: the role of Sp1.J Cell Mol Med. 2015 Mar;19(3):630-41. doi: 10.1111/jcmm.12476. Epub 2015 Jan 19. J Cell Mol Med. 2015. PMID: 25598321 Free PMC article.
-
HOTAIR-mediated reciprocal regulation of EZH2 and DNMT1 contribute to polyphyllin I-inhibited growth of castration-resistant prostate cancer cells in vitro and in vivo.Biochim Biophys Acta Gen Subj. 2018 Mar;1862(3):589-599. doi: 10.1016/j.bbagen.2017.12.001. Epub 2017 Dec 6. Biochim Biophys Acta Gen Subj. 2018. PMID: 29221985
-
Repression of PDK1- and LncRNA HOTAIR-Mediated EZH2 Gene Expression Contributes to the Enhancement of Atractylenolide 1 and Erlotinib in the Inhibition of Human Lung Cancer Cells.Cell Physiol Biochem. 2018;49(4):1615-1632. doi: 10.1159/000493497. Epub 2018 Sep 17. Cell Physiol Biochem. 2018. PMID: 30223276
-
β-elemene: A promising natural compound in lung cancer therapy.Eur J Pharmacol. 2025 Jun 15;997:177399. doi: 10.1016/j.ejphar.2025.177399. Epub 2025 Mar 8. Eur J Pharmacol. 2025. PMID: 40064226 Review.
Cited by
-
Natural Bioactive Compounds Targeting Epigenetic Pathways in Cancer: A Review on Alkaloids, Terpenoids, Quinones, and Isothiocyanates.Nutrients. 2021 Oct 22;13(11):3714. doi: 10.3390/nu13113714. Nutrients. 2021. PMID: 34835969 Free PMC article. Review.
-
DNMT1 modulates interneuron morphology by regulating Pak6 expression through crosstalk with histone modifications.Epigenetics. 2018;13(5):536-556. doi: 10.1080/15592294.2018.1475980. Epub 2018 Aug 7. Epigenetics. 2018. PMID: 29912614 Free PMC article.
-
β-elemene Isopropanolamine Derivative LXX-8250 Induces Apoptosis Through Impairing Autophagic Flux via PFKFB4 Repression in Melanoma Cells.Front Pharmacol. 2022 Aug 10;13:900973. doi: 10.3389/fphar.2022.900973. eCollection 2022. Front Pharmacol. 2022. PMID: 36034839 Free PMC article.
-
FBP1 induced by β-elemene enhances the sensitivity of gefitinib in lung cancer.Thorac Cancer. 2023 Feb;14(4):371-380. doi: 10.1111/1759-7714.14750. Epub 2022 Dec 16. Thorac Cancer. 2023. PMID: 36525508 Free PMC article.
-
β-Elemene alleviates cisplatin resistance in oral squamous cell carcinoma cell via inhibiting JAK2/STAT3 pathway in vitro and in vivo.Cancer Cell Int. 2022 Jul 31;22(1):244. doi: 10.1186/s12935-022-02650-7. Cancer Cell Int. 2022. PMID: 35909161 Free PMC article.
References
-
- Hildesheim A, Wang CP. Genetic predisposition factors and nasopharyngeal carcinoma risk: a review of epidemiological association studies, 2000-2011: Rosetta Stone for NPC: genetics, viral infection, and other environmental factors. Semin Cancer Biol. 2012;22:107–116. doi: 10.1016/j.semcancer.2012.01.007. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
