

Rapid breeding of parthenocarpic tomato plants using CRISPR/Cas9
- PMID: 28360425
- PMCID: PMC5428692
- DOI: 10.1038/s41598-017-00501-4
Rapid breeding of parthenocarpic tomato plants using CRISPR/Cas9
Erratum in
-
Author Correction: Rapid breeding of parthenocarpic tomato plants using CRISPR/Cas9.Sci Rep. 2020 Oct 2;10(1):16776. doi: 10.1038/s41598-020-71765-6. Sci Rep. 2020. PMID: 33009418 Free PMC article.
Abstract
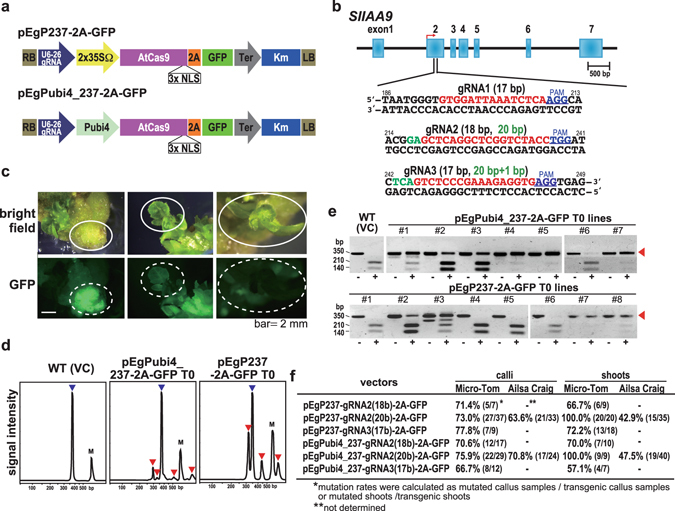
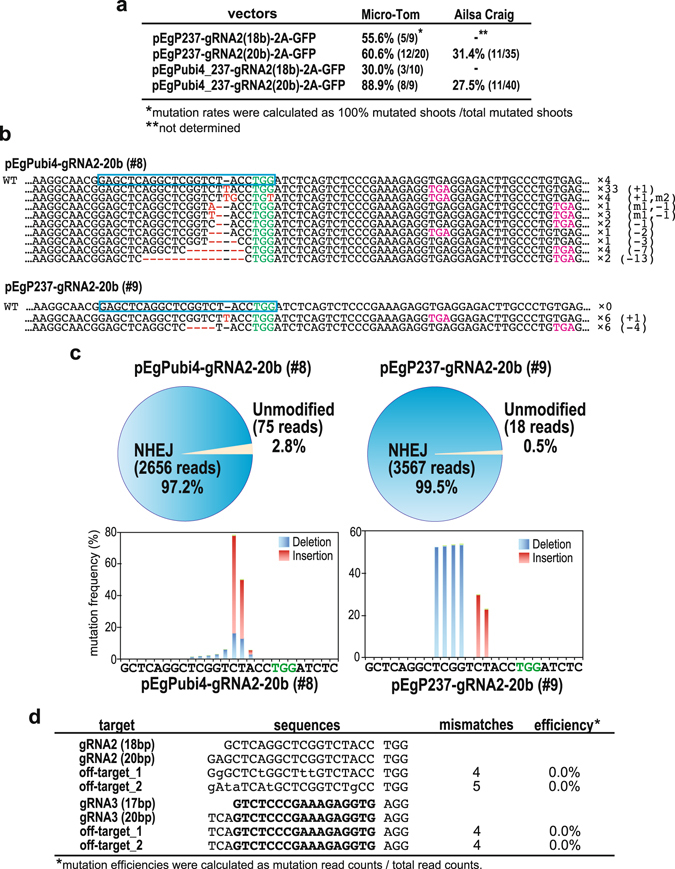
Parthenocarpy in horticultural crop plants is an important trait with agricultural value for various industrial purposes as well as direct eating quality. Here, we demonstrate a breeding strategy to generate parthenocarpic tomato plants using the CRISPR/Cas9 system. We optimized the CRISPR/Cas9 system to introduce somatic mutations effectively into SlIAA9-a key gene controlling parthenocarpy-with mutation rates of up to 100% in the T0 generation. Furthermore, analysis of off-target mutations using deep sequencing indicated that our customized gRNAs induced no additional mutations in the host genome. Regenerated mutants exhibited morphological changes in leaf shape and seedless fruit-a characteristic of parthenocarpic tomato. And the segregated next generation (T1) also showed a severe phenotype associated with the homozygous mutated genome. The system developed here could be applied to produce parthenocarpic tomato in a wide variety of cultivars, as well as other major horticultural crops, using this precise and rapid breeding technique.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

Similar articles
-
Advances in application of genome editing in tomato and recent development of genome editing technology.Theor Appl Genet. 2021 Sep;134(9):2727-2747. doi: 10.1007/s00122-021-03874-3. Epub 2021 Jun 2. Theor Appl Genet. 2021. PMID: 34076729 Free PMC article. Review.
-
CRISPR/Cas9-mediated efficient and heritable targeted mutagenesis in tomato plants in the first and later generations.Sci Rep. 2016 Apr 21;6:24765. doi: 10.1038/srep24765. Sci Rep. 2016. PMID: 27097775 Free PMC article.
-
The parthenocarpic gene Pat-k is generated by a natural mutation of SlAGL6 affecting fruit development in tomato (Solanum lycopersicum L.).BMC Plant Biol. 2018 Apr 27;18(1):72. doi: 10.1186/s12870-018-1285-6. BMC Plant Biol. 2018. PMID: 29699487 Free PMC article.
-
Tomato facultative parthenocarpy results from SlAGAMOUS-LIKE 6 loss of function.Plant Biotechnol J. 2017 May;15(5):634-647. doi: 10.1111/pbi.12662. Epub 2016 Dec 27. Plant Biotechnol J. 2017. PMID: 27862876 Free PMC article.
-
A status-quo review on CRISPR-Cas9 gene editing applications in tomato.Int J Biol Macromol. 2021 Nov 1;190:120-129. doi: 10.1016/j.ijbiomac.2021.08.169. Epub 2021 Aug 30. Int J Biol Macromol. 2021. PMID: 34474054 Review.
Cited by
-
Advances in application of genome editing in tomato and recent development of genome editing technology.Theor Appl Genet. 2021 Sep;134(9):2727-2747. doi: 10.1007/s00122-021-03874-3. Epub 2021 Jun 2. Theor Appl Genet. 2021. PMID: 34076729 Free PMC article. Review.
-
Applications of CRISPR/Cas genome editing in economically important fruit crops: recent advances and future directions.Mol Hortic. 2023 Jan 28;3(1):1. doi: 10.1186/s43897-023-00049-0. Mol Hortic. 2023. PMID: 37789479 Free PMC article. Review.
-
Advancing vegetable genetics with gene editing: a pathway to food security and nutritional resilience in climate-shifted environments.Funct Integr Genomics. 2025 Feb 1;25(1):31. doi: 10.1007/s10142-025-01533-0. Funct Integr Genomics. 2025. PMID: 39891757 Review.
-
Genomic, Transcriptomic and Epigenomic Tools to Study the Domestication of Plants and Animals: A Field Guide for Beginners.Front Genet. 2020 Jul 15;11:742. doi: 10.3389/fgene.2020.00742. eCollection 2020. Front Genet. 2020. PMID: 32760427 Free PMC article. Review.
-
Efficient Multiplex Genome Editing Induces Precise, and Self-Ligated Type Mutations in Tomato Plants.Front Plant Sci. 2018 Jul 3;9:916. doi: 10.3389/fpls.2018.00916. eCollection 2018. Front Plant Sci. 2018. PMID: 30018630 Free PMC article.
References
-
- Srivastava A, Handa A. Hormonal regulation of tomato fruit development: a molecular perspective. J. Plant Growth Regul. 2005;24:67–82. doi: 10.1007/s00344-005-0015-0. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
