

Adaptation Mechanisms in the Evolution of Moss Defenses to Microbes
- PMID: 28360923
- PMCID: PMC5350094
- DOI: 10.3389/fpls.2017.00366
Adaptation Mechanisms in the Evolution of Moss Defenses to Microbes
Abstract
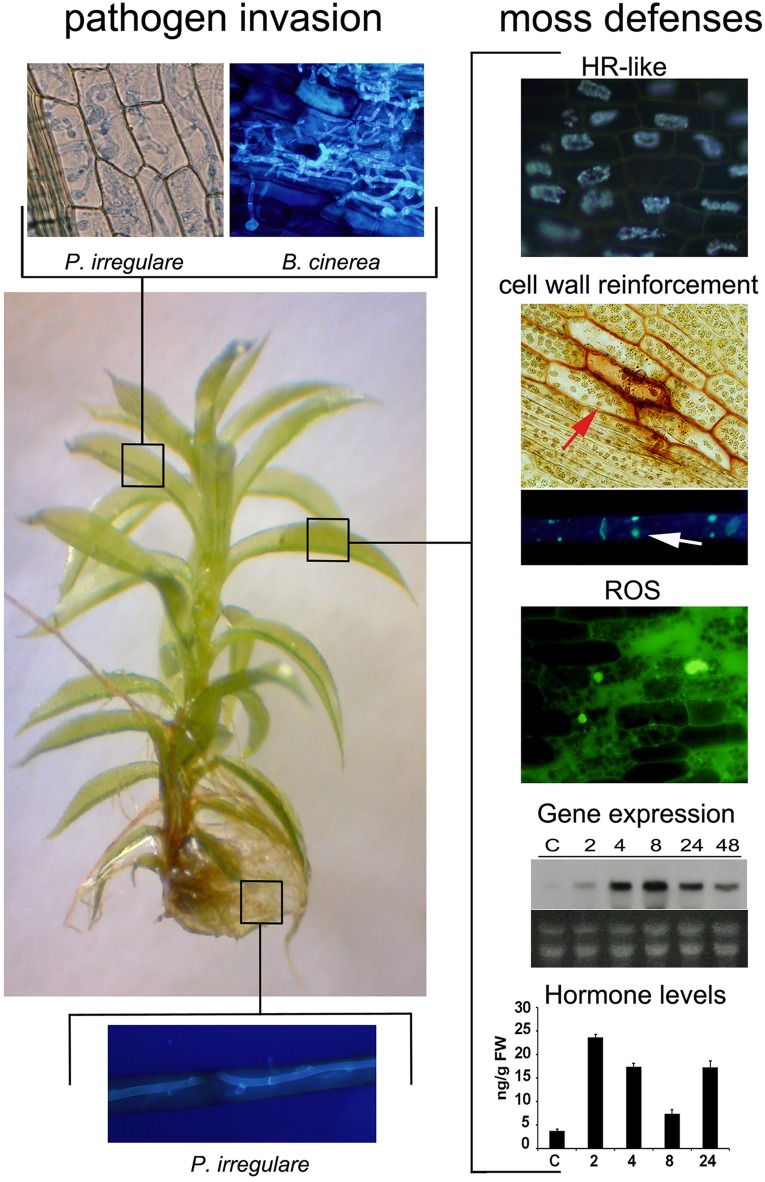
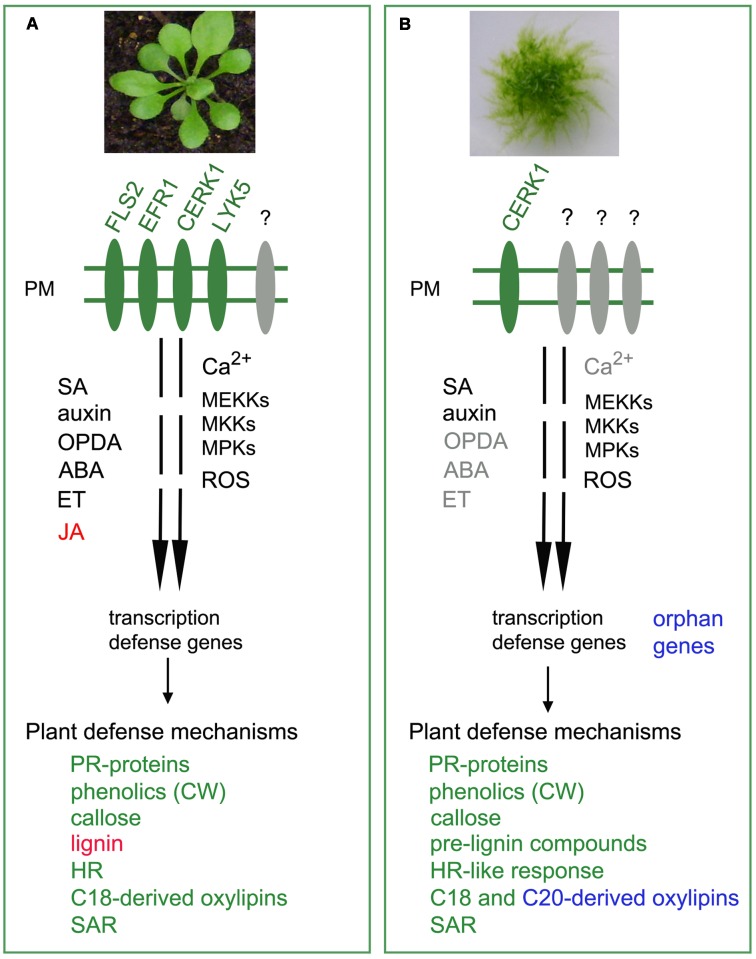
Bryophytes, including mosses, liverworts and hornworts are early land plants that have evolved key adaptation mechanisms to cope with abiotic stresses and microorganisms. Microbial symbioses facilitated plant colonization of land by enhancing nutrient uptake leading to improved plant growth and fitness. In addition, early land plants acquired novel defense mechanisms to protect plant tissues from pre-existing microbial pathogens. Due to its evolutionary stage linking unicellular green algae to vascular plants, the non-vascular moss Physcomitrella patens is an interesting organism to explore the adaptation mechanisms developed in the evolution of plant defenses to microbes. Cellular and biochemical approaches, gene expression profiles, and functional analysis of genes by targeted gene disruption have revealed that several defense mechanisms against microbial pathogens are conserved between mosses and flowering plants. P. patens perceives pathogen associated molecular patterns by plasma membrane receptor(s) and transduces the signal through a MAP kinase (MAPK) cascade leading to the activation of cell wall associated defenses and expression of genes that encode proteins with different roles in plant resistance. After pathogen assault, P. patens also activates the production of ROS, induces a HR-like reaction and increases levels of some hormones. Furthermore, alternative metabolic pathways are present in P. patens leading to the production of a distinct metabolic scenario than flowering plants that could contribute to defense. P. patens has acquired genes by horizontal transfer from prokaryotes and fungi, and some of them could represent adaptive benefits for resistance to biotic stress. In this review, the current knowledge related to the evolution of plant defense responses against pathogens will be discussed, focusing on the latest advances made in the model plant P. patens.
Keywords: adaptation mechanisms; evolution; horizontal gene transfer; moss-microbe interactions; pathogens; plant defenses.
Figures
Similar articles
-
Transcriptional profiling reveals conserved and species-specific plant defense responses during the interaction of Physcomitrium patens with Botrytis cinerea.Plant Mol Biol. 2021 Nov;107(4-5):365-385. doi: 10.1007/s11103-021-01116-0. Epub 2021 Feb 1. Plant Mol Biol. 2021. PMID: 33521880
-
Activation of Defense Mechanisms against Pathogens in Mosses and Flowering Plants.Int J Mol Sci. 2013 Feb 4;14(2):3178-200. doi: 10.3390/ijms14023178. Int J Mol Sci. 2013. PMID: 23380962 Free PMC article.
-
The Moss Physcomitrella patens as a Model System to Study Interactions between Plants and Phytopathogenic Fungi and Oomycetes.J Pathog. 2011;2011:719873. doi: 10.4061/2011/719873. Epub 2011 Aug 4. J Pathog. 2011. PMID: 22567339 Free PMC article.
-
Moss transcription factors regulating development and defense responses to stress.J Exp Bot. 2022 Jul 16;73(13):4546-4561. doi: 10.1093/jxb/erac055. J Exp Bot. 2022. PMID: 35167679 Review.
-
Leaf Morphogenesis: Insights From the Moss Physcomitrium patens.Front Plant Sci. 2021 Sep 23;12:736212. doi: 10.3389/fpls.2021.736212. eCollection 2021. Front Plant Sci. 2021. PMID: 34630486 Free PMC article. Review.
Cited by
-
Beyond Photoprotection: The Multifarious Roles of Flavonoids in Plant Terrestrialization.Int J Mol Sci. 2022 May 9;23(9):5284. doi: 10.3390/ijms23095284. Int J Mol Sci. 2022. PMID: 35563675 Free PMC article. Review.
-
Great chemistry between us: The link between plant chemical defenses and butterfly evolution.Ecol Evol. 2021 May 27;11(13):8595-8613. doi: 10.1002/ece3.7673. eCollection 2021 Jul. Ecol Evol. 2021. PMID: 34257918 Free PMC article.
-
An Overview of PRR- and NLR-Mediated Immunities: Conserved Signaling Components across the Plant Kingdom That Communicate Both Pathways.Int J Mol Sci. 2022 Oct 26;23(21):12974. doi: 10.3390/ijms232112974. Int J Mol Sci. 2022. PMID: 36361764 Free PMC article. Review.
-
On plant defense signaling networks and early land plant evolution.Commun Integr Biol. 2018 Aug 9;11(3):1-14. doi: 10.1080/19420889.2018.1486168. eCollection 2018. Commun Integr Biol. 2018. PMID: 30214675 Free PMC article. Review.
-
Gene refashioning through innovative shifting of reading frames in mosses.Nat Commun. 2018 Apr 19;9(1):1555. doi: 10.1038/s41467-018-04025-x. Nat Commun. 2018. PMID: 29674719 Free PMC article.
References
-
- Adie B. A., Pérez-Pérez J., Pérez-Pérez M. M., Godoy M., Sánchez-Serrano J. J., Schmelz E. A., et al. (2007). ABA is an essential signal for plant resistance to pathogens affecting JA biosynthesis and the activation of defenses in Arabidopsis. Plant Cell 19 1665–1681. 10.1105/tpc.106.048041 - DOI - PMC - PubMed
-
- Andersson R. A., Akita M., Pirhonen M., Gammelgård E., Valkonen J. P. T. (2005). Moss-Erwinia pathosystem reveals possible similarities in pathogenesis and pathogen defense in vascular and nonvascular plants. J. Gen. Plant Pathol. 71 23–28. 10.1007/s10327-004-0154-3 - DOI
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources