

Role of primary cilia in non-dividing and post-mitotic cells
- PMID: 28361305
- PMCID: PMC5487853
- DOI: 10.1007/s00441-017-2599-7
Role of primary cilia in non-dividing and post-mitotic cells
Abstract
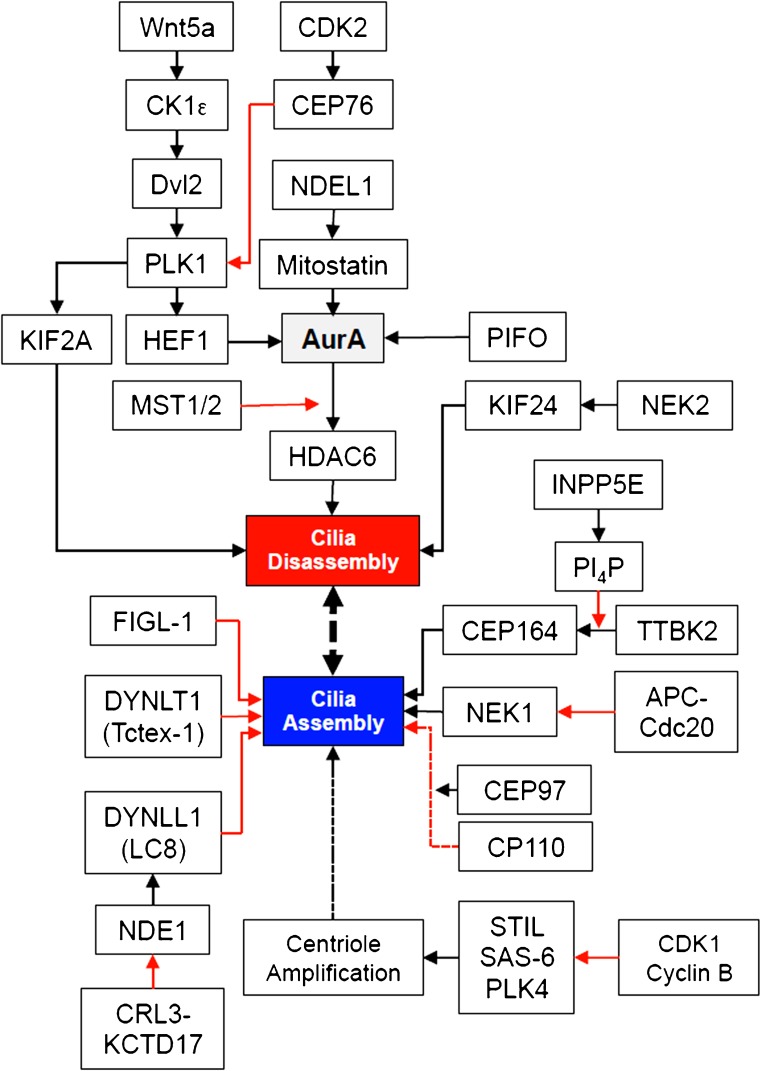
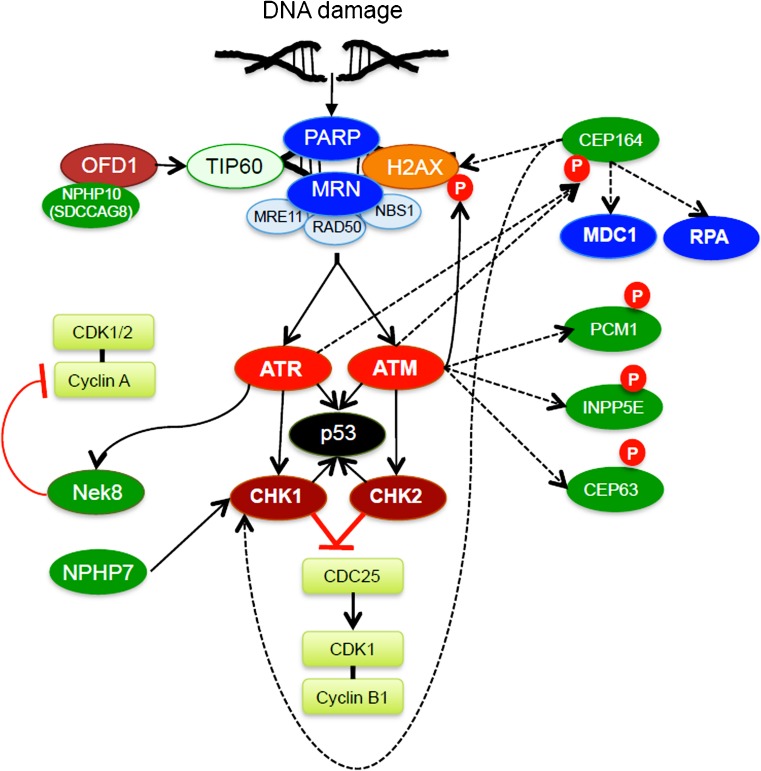
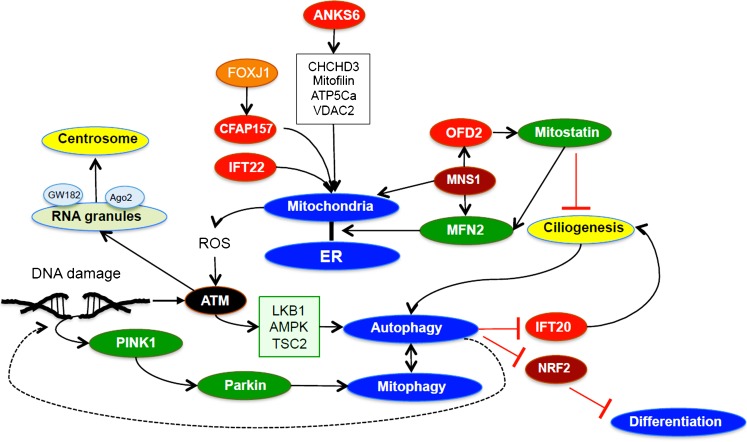
The essential role of primary (non-motile) cilia during the development of multi-cellular tissues and organs is well established and is underlined by severe disease manifestations caused by mutations in cilia-associated molecules that are collectively termed ciliopathies. However, the role of primary cilia in non-dividing and terminally differentiated, post-mitotic cells is less well understood. Although the prevention of cells from re-entering the cell cycle may represent a major chore, primary cilia have recently been linked to DNA damage responses, autophagy and mitochondria. Given this connectivity, primary cilia in non-dividing cells are well positioned to form a signaling hub outside of the nucleus. Such a center could integrate information to initiate responses and to maintain cellular homeostasis if cell survival is jeopardized. These more discrete functions may remain undetected until differentiated cells are confronted with emergencies.
Keywords: Autophagy; Ciliopathy; DNA damage response; Mitochondria; mTOR.
Conflict of interest statement
The author declares that he has no conflict of interest.
Figures
Similar articles
-
Primary cilia and the DNA damage response: linking a cellular antenna and nuclear signals.Biochem Soc Trans. 2021 Apr 30;49(2):829-841. doi: 10.1042/BST20200751. Biochem Soc Trans. 2021. PMID: 33843966 Review.
-
The Nuclear Arsenal of Cilia.Dev Cell. 2019 Apr 22;49(2):161-170. doi: 10.1016/j.devcel.2019.03.009. Dev Cell. 2019. PMID: 31014478 Review.
-
Effects of Sesquiterpene Lactones on Primary Cilia Formation (Ciliogenesis).Toxins (Basel). 2023 Oct 27;15(11):632. doi: 10.3390/toxins15110632. Toxins (Basel). 2023. PMID: 37999495 Free PMC article.
-
Crosstalk between cilia and autophagy: implication for human diseases.Autophagy. 2023 Jan;19(1):24-43. doi: 10.1080/15548627.2022.2067383. Epub 2022 May 25. Autophagy. 2023. PMID: 35613303 Free PMC article. Review.
-
The inner junction protein CFAP20 functions in motile and non-motile cilia and is critical for vision.Nat Commun. 2022 Nov 3;13(1):6595. doi: 10.1038/s41467-022-33820-w. Nat Commun. 2022. PMID: 36329026 Free PMC article.
Cited by
-
Mucociliary Respiratory Epithelium Integrity in Molecular Defense and Susceptibility to Pulmonary Viral Infections.Biology (Basel). 2021 Jan 29;10(2):95. doi: 10.3390/biology10020095. Biology (Basel). 2021. PMID: 33572760 Free PMC article. Review.
-
Artificial Intelligence Approaches to Assessing Primary Cilia.J Vis Exp. 2021 May 1;(171):10.3791/62521. doi: 10.3791/62521. J Vis Exp. 2021. PMID: 33999029 Free PMC article.
-
Co-expression of xenopsin and rhabdomeric opsin in photoreceptors bearing microvilli and cilia.Elife. 2017 Sep 6;6:e23435. doi: 10.7554/eLife.23435. Elife. 2017. PMID: 28876222 Free PMC article.
-
Mutations in NEK1 cause ciliary dysfunction as a novel pathogenic mechanism in amyotrophic lateral sclerosis.Mol Neurodegener. 2025 May 20;20(1):59. doi: 10.1186/s13024-025-00848-7. Mol Neurodegener. 2025. PMID: 40389989 Free PMC article.
-
Analysis from the perspective of cilia: the protective effect of PARP inhibitors on visual function during light-induced damage.Int Ophthalmol. 2020 Apr;40(4):1017-1027. doi: 10.1007/s10792-019-01245-y. Epub 2019 Dec 4. Int Ophthalmol. 2020. PMID: 31802371
References
-
- Abramowicz I, Carpenter G, Alfieri M, Colnaghi R, Outwin E, Parent P, Thauvin-Robinet C, Iaconis D, Franco B, O’Driscoll M (2016) Oral-facial-digital syndrome type I cells exhibit impaired DNA repair; unanticipated consequences of defective OFD1 outside of the cilia network. Hum Mol Genet 2016:ddw364. 10.1093/hmg10.1093/hmg - PubMed
-
- Airik R, Slaats GG, Guo Z, Weiss AC, Khan N, Ghosh A, Hurd TW, Bekker-Jensen S, Schroder JM, Elledge SJ, Andersen JS, Kispert A, Castelli M, Boletta A, Giles RH, Hildebrandt F. Renal-retinal ciliopathy gene Sdccag8 regulates DNA damage response signaling. J Am Soc Nephrol. 2014;25:2573–2583. doi: 10.1681/ASN.2013050565. - DOI - PMC - PubMed
-
- Alexander A, Cai SL, Kim J, Nanez A, Sahin M, MacLean KH, Inoki K, Guan KL, Shen J, Person MD, Kusewitt D, Mills GB, Kastan MB, Walker CL. ATM signals to TSC2 in the cytoplasm to regulate mTORC1 in response to ROS. Proc Natl Acad Sci U S A. 2010;107:4153–4158. doi: 10.1073/pnas.0913860107. - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
