

Development of the macaque face-patch system
- PMID: 28361890
- PMCID: PMC5381009
- DOI: 10.1038/ncomms14897
Development of the macaque face-patch system
Abstract
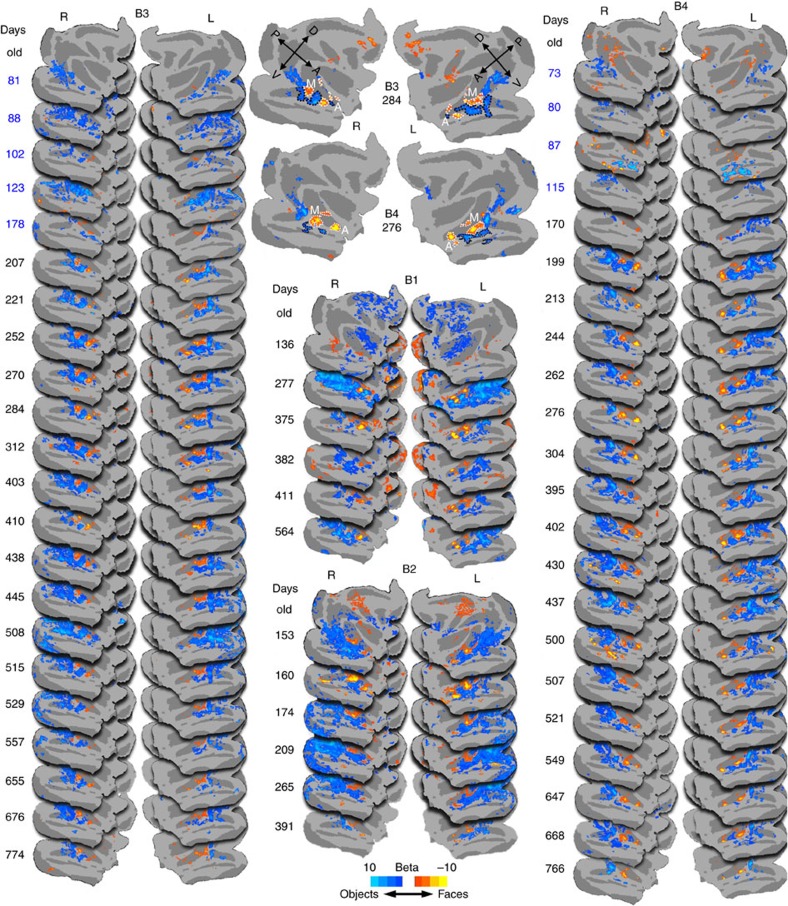
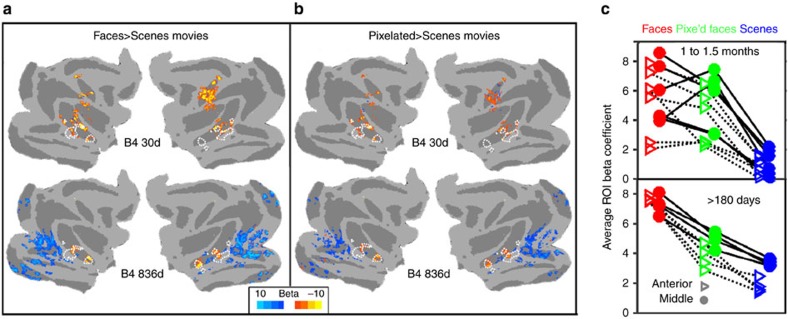
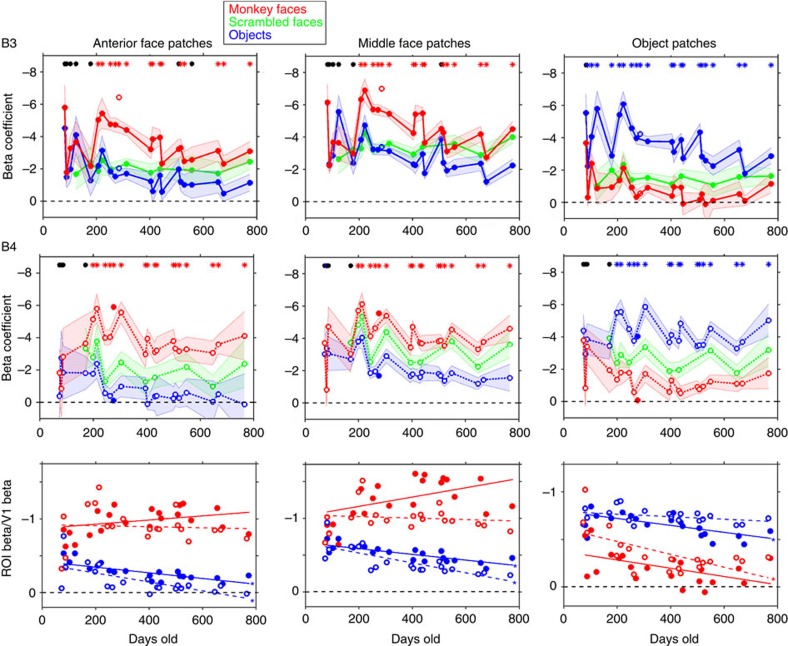
Face recognition is highly proficient in humans and other social primates; it emerges in infancy, but the development of the neural mechanisms supporting this behaviour is largely unknown. We use blood-volume functional MRI to monitor longitudinally the responsiveness to faces, scrambled faces, and objects in macaque inferotemporal cortex (IT) from 1 month to 2 years of age. During this time selective responsiveness to monkey faces emerges. Some functional organization is present at 1 month; face-selective patches emerge over the first year of development, and are remarkably stable once they emerge. Face selectivity is refined by a decreasing responsiveness to non-face stimuli.
Conflict of interest statement
The authors declare no competing financial interests.
Figures




References
-
- Chan P., Downing M. E. & Kanwisher N. Domain specificity in visual cortex. Cereb. Cortex 16, 1453–1461 (2006). - PubMed
-
- McKone E., Crookes K., Jeffery L. & Dilks D. D. A critical review of the development of face recognition: experience is less important than previously believed. Cogn. Neuropsychol. 29, 174–212 (2012). - PubMed
-
- Hasson U., Levy I., Behrmann M., Hendler T. & Malach R. Eccentricity bias as an organizing principle for human high-order object areas. Neuron 34, 479–490 (2002). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
