

Single-cell RNA sequencing identifies distinct mouse medial ganglionic eminence cell types
- PMID: 28361918
- PMCID: PMC5374502
- DOI: 10.1038/srep45656
Single-cell RNA sequencing identifies distinct mouse medial ganglionic eminence cell types
Abstract
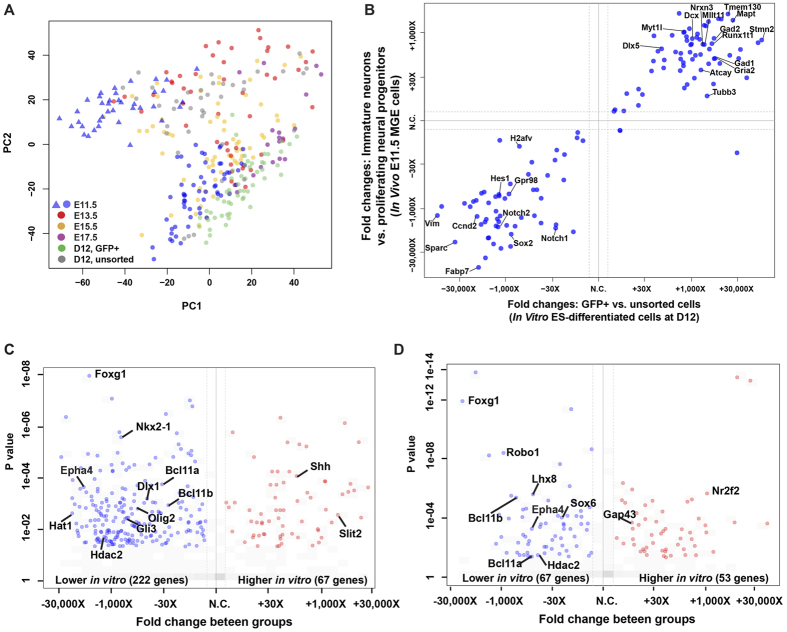
Many subtypes of cortical interneurons (CINs) are found in adult mouse cortices, but the mechanism generating their diversity remains elusive. We performed single-cell RNA sequencing on the mouse embryonic medial ganglionic eminence (MGE), the major birthplace for CINs, and on MGE-like cells differentiated from embryonic stem cells. Two distinct cell types were identified as proliferating neural progenitors and immature neurons, both of which comprised sub-populations. Although lineage development of MGE progenitors was reconstructed and immature neurons were characterized as GABAergic, cells that might correspond to precursors of different CINs were not identified. A few non-neuronal cell types were detected, including microglia. In vitro MGE-like cells resembled bona fide MGE cells but expressed lower levels of Foxg1 and Epha4. Together, our data provide detailed understanding of the embryonic MGE developmental program and suggest how CINs are specified.
Conflict of interest statement
All authors except for J.L.R. are/were employees of Genentech and declare no conflicts of interest. J.L.R. is supported by R01 Grant MH081880 from NIMH and is a Founder and Scientific Advisory Board of Neurona, Inc.
Figures
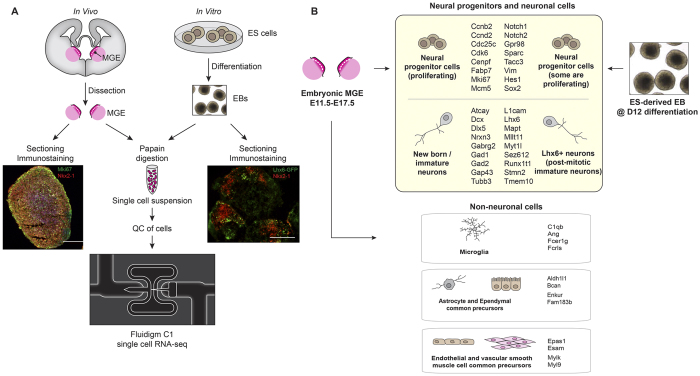
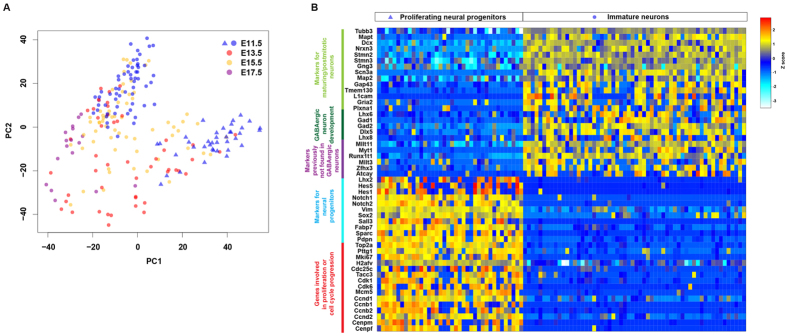
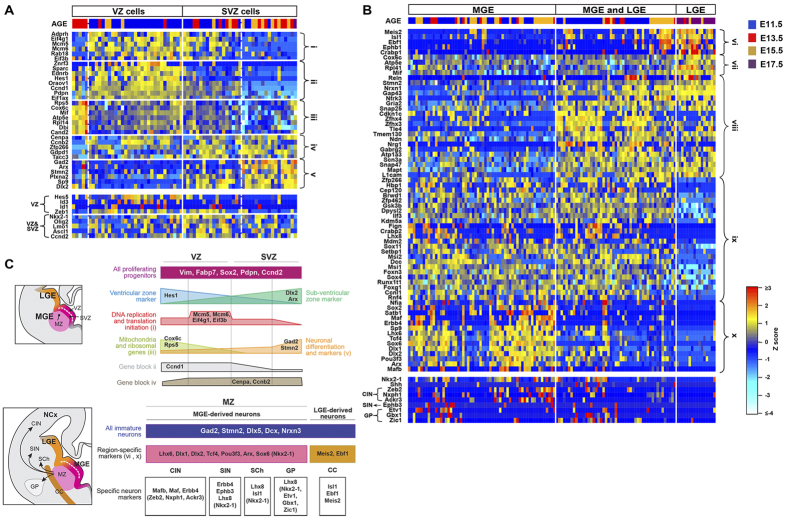
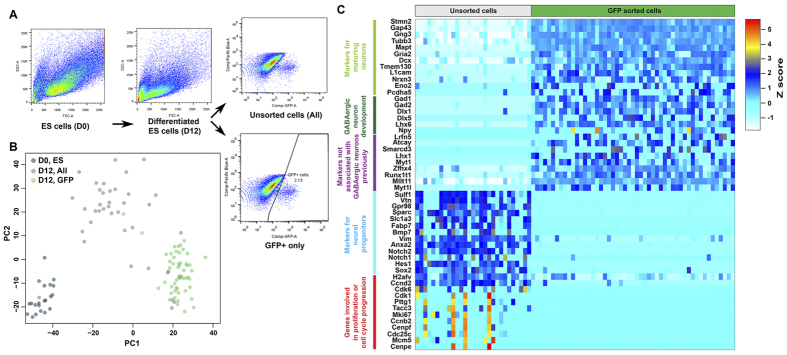
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
