

Characterization of signal bias at the GLP-1 receptor induced by backbone modification of GLP-1
- PMID: 28363772
- PMCID: PMC5794489
- DOI: 10.1016/j.bcp.2017.03.018
Characterization of signal bias at the GLP-1 receptor induced by backbone modification of GLP-1
Abstract
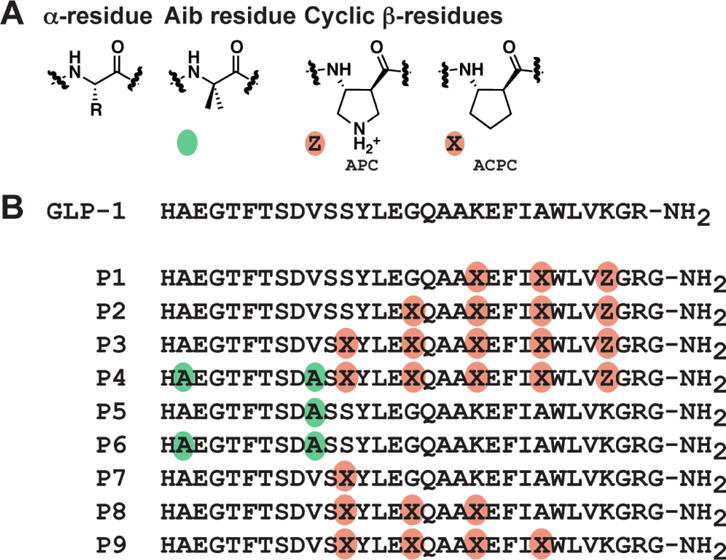
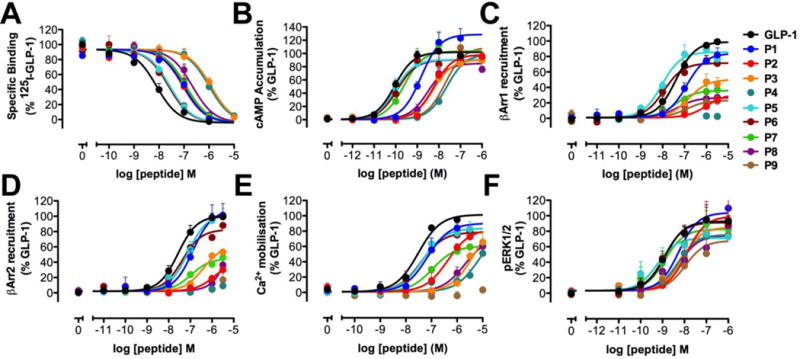
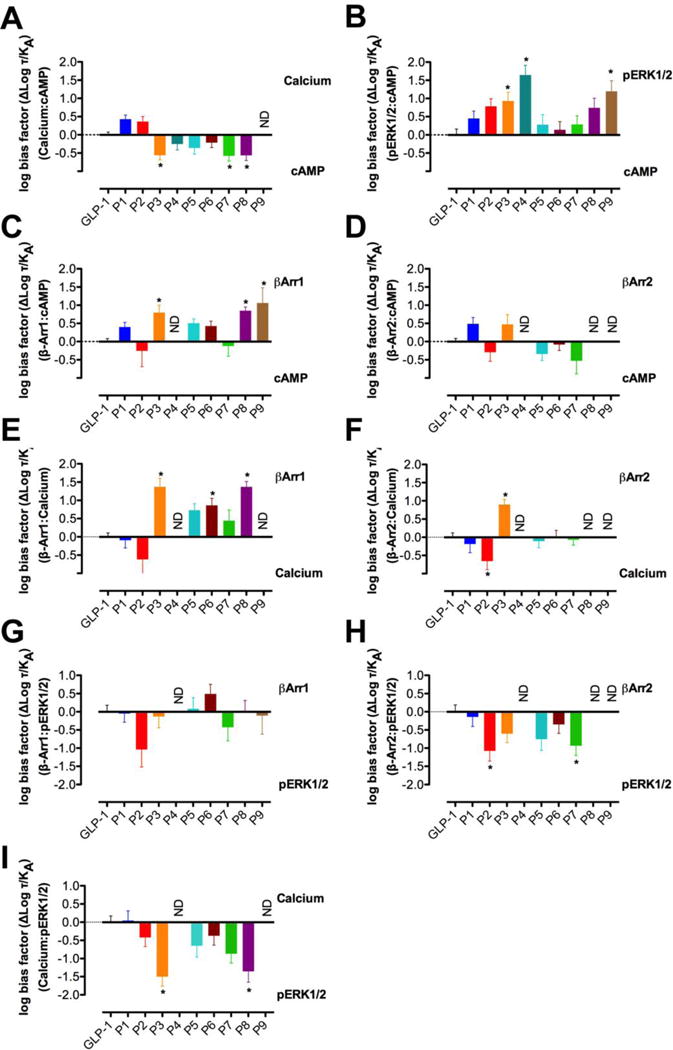
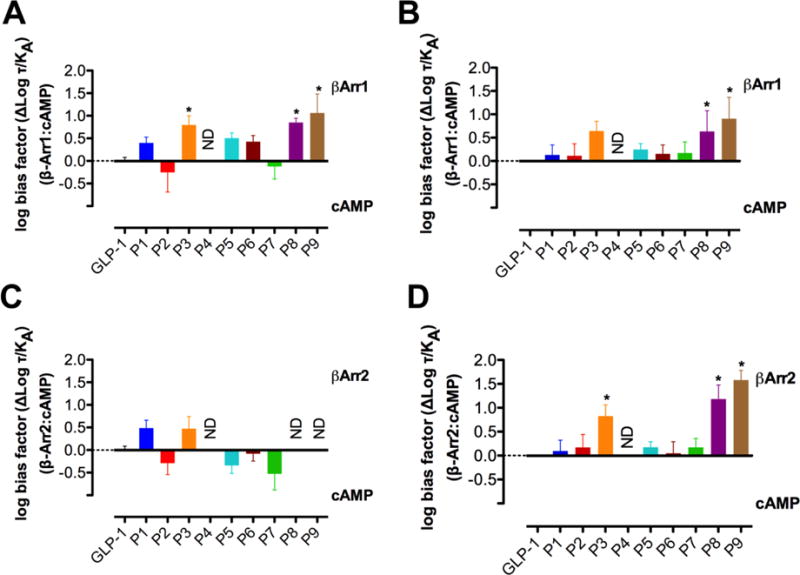
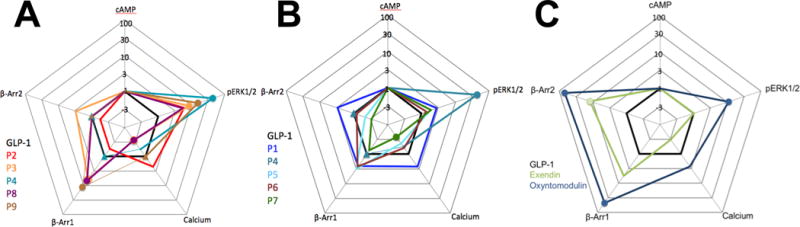
The glucagon-like peptide-1 receptor (GLP-1R) is a class B G protein-coupled receptor that is a major therapeutic target for the treatment of type 2 diabetes. Activation of this receptor promotes insulin secretion and blood glucose regulation. The GLP-1R can initiate signaling through several intracellular pathways upon activation by GLP-1. GLP-1R ligands that preferentially stimulate subsets among the natural signaling pathways ("biased agonists") could be useful as tools for elucidating the consequences of specific pathways and might engender therapeutic agents with tailored effects. Using HEK-293 cells recombinantly expressing human GLP-1R, we have previously reported that backbone modification of GLP-1, via replacement of selected α-amino acid residues with β-amino acid residues, generates GLP-1 analogues with distinctive preferences for promoting G protein activation versus β-arrestin recruitment. Here, we have explored the influence of cell background across these two parameters and expanded our analysis to include affinity and other key signaling pathways (intracellular calcium mobilization and ERK phosphorylation) using recombinant human GLP-1R expressed in a CHO cell background, which has been used extensively to demonstrate biased agonism of GLP-1R ligands. The new data indicate that α/β-peptide analogues of GLP-1 exhibit a range of distinct bias profiles relative to GLP-1 and that broad assessment of signaling endpoints is required to reveal the spectrum of behavior of modified peptides. These results support the view that backbone modification via α→β amino acid replacement can enable rapid discovery of peptide hormone analogues that display substantial signal bias at a cognate GPCR.
Keywords: Biased agonism; Cell signaling; G protein coupled receptor; Glucagon-like peptide-1 receptor; Peptides.
Copyright © 2017 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare the following competing financial interest(s): S.H.G. is an inventor on a patent application covering GLP-1 analogues described here; S.H.G. is a cofounder of Longevity Biotech, Inc., which is pursuing biomedical applications of α/β-peptides.
Figures
References
-
- Zaccardi F, Webb DR, Yates T, Davies MJ. Pathophysiology of type 1 and type 2 diabetes mellitus: a 90-year perspective. Postgrad Med J. 2016;92:63–69. - PubMed
-
- DeFronzo RA. Pathogensis of Type 2 (non-insulin dependent) diabetes mellitus: a balanced overview. Diabetologia. 1992;35:389–397. - PubMed
-
- Hundal RS, Inzucchi SE. Metformin: new understandings, new uses. Drugs. 2003;63:1879–1894. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
