

Structure-Based Engineering of an Artificially Generated NADP+-Dependent d-Amino Acid Dehydrogenase
- PMID: 28363957
- PMCID: PMC5440713
- DOI: 10.1128/AEM.00491-17
Structure-Based Engineering of an Artificially Generated NADP+-Dependent d-Amino Acid Dehydrogenase
Abstract
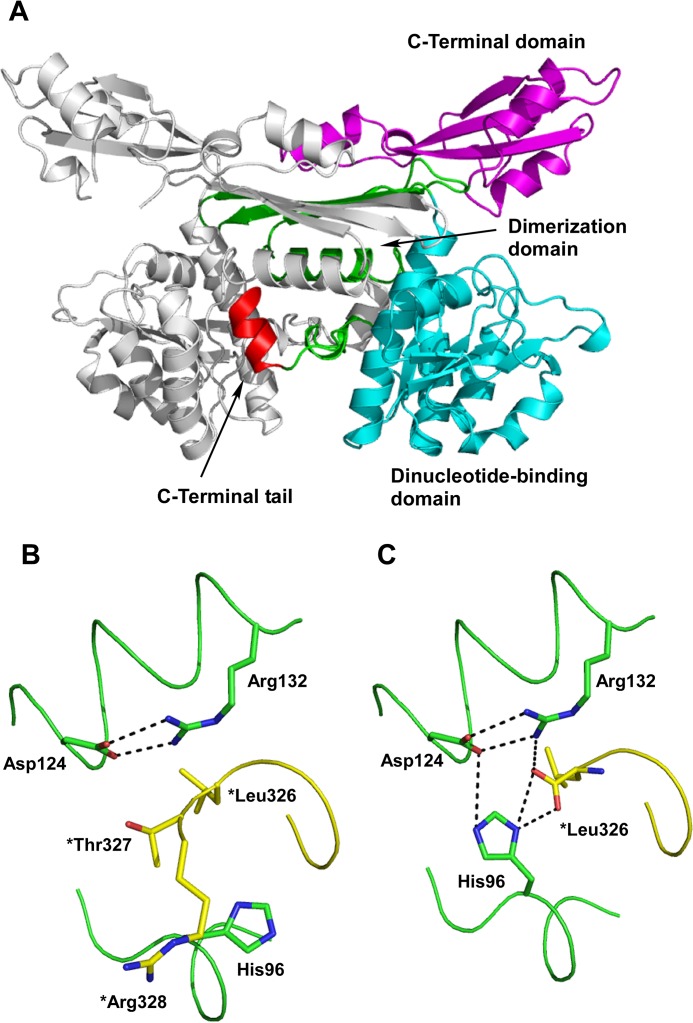
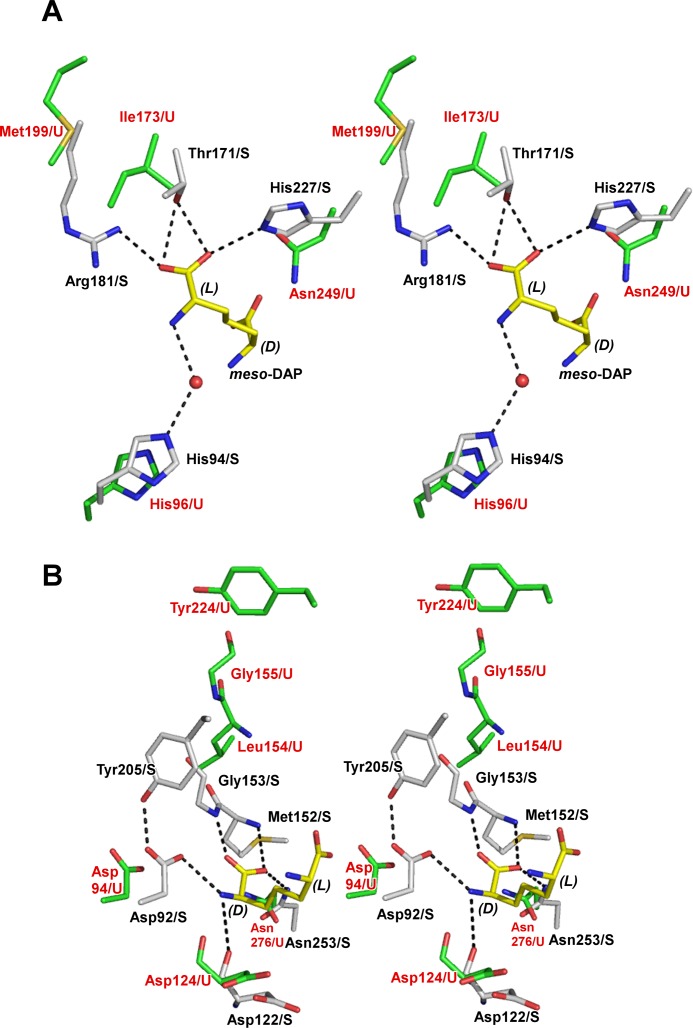
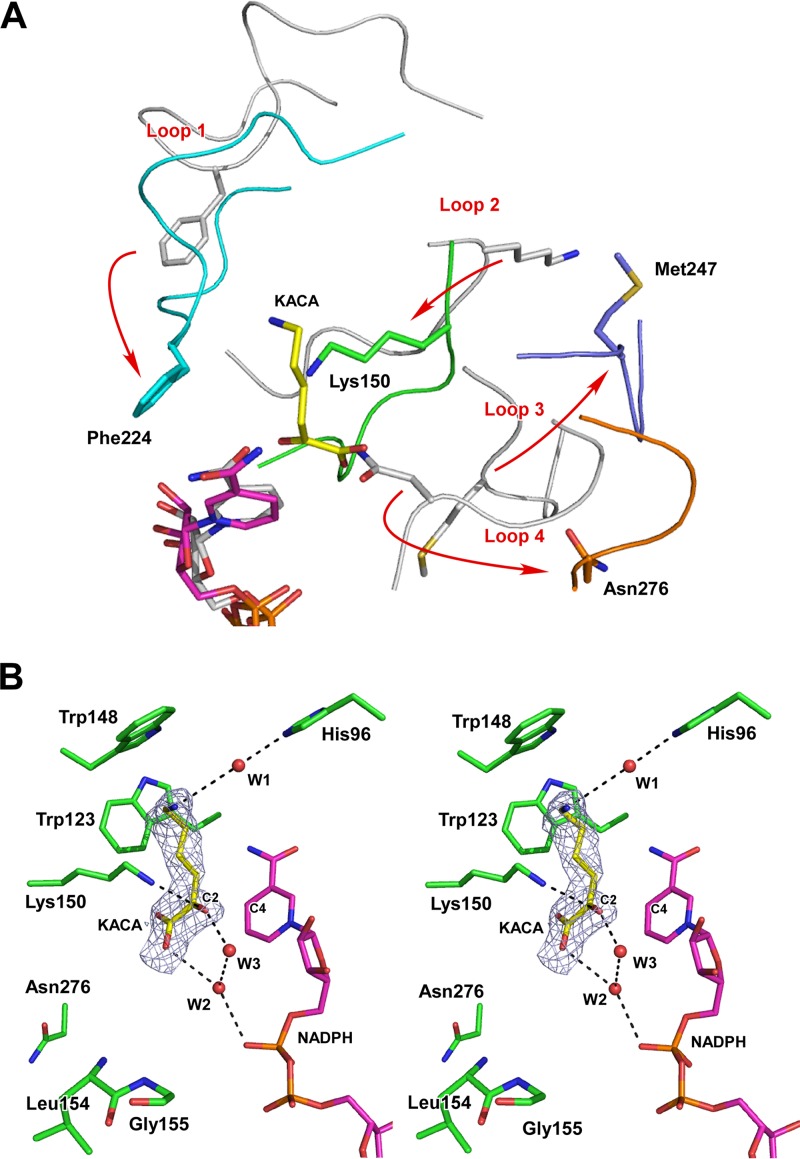
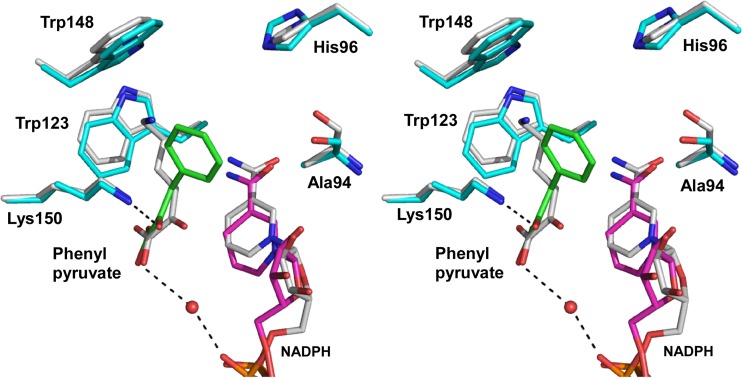
A stable NADP+-dependent d-amino acid dehydrogenase (DAADH) was recently created from Ureibacillus thermosphaericusmeso-diaminopimelate dehydrogenase through site-directed mutagenesis. To produce a novel DAADH mutant with different substrate specificity, the crystal structure of apo-DAADH was determined at a resolution of 1.78 Å, and the amino acid residues responsible for the substrate specificity were evaluated using additional site-directed mutagenesis. By introducing a single D94A mutation, the enzyme's substrate specificity was dramatically altered; the mutant utilized d-phenylalanine as the most preferable substrate for oxidative deamination and had a specific activity of 5.33 μmol/min/mg at 50°C, which was 54-fold higher than that of the parent DAADH. In addition, the specific activities of the mutant toward d-leucine, d-norleucine, d-methionine, d-isoleucine, and d-tryptophan were much higher (6 to 25 times) than those of the parent enzyme. For reductive amination, the D94A mutant exhibited extremely high specific activity with phenylpyruvate (16.1 μmol/min/mg at 50°C). The structures of the D94A-Y224F double mutant in complex with NADP+ and in complex with both NADPH and 2-keto-6-aminocapronic acid (lysine oxo-analogue) were then determined at resolutions of 1.59 Å and 1.74 Å, respectively. The phenylpyruvate-binding model suggests that the D94A mutation prevents the substrate phenyl group from sterically clashing with the side chain of Asp94. A structural comparison suggests that both the enlarged substrate-binding pocket and enhanced hydrophobicity of the pocket are mainly responsible for the high reactivity of the D94A mutant toward the hydrophobic d-amino acids with bulky side chains.IMPORTANCE In recent years, the potential uses for d-amino acids as source materials for the industrial production of medicines, seasonings, and agrochemicals have been growing. To date, several methods have been used for the production of d-amino acids, but all include tedious steps. The use of NAD(P)+-dependent d-amino acid dehydrogenase (DAADH) makes single-step production of d-amino acids from oxo-acid analogs and ammonia possible. We recently succeeded in creating a stable DAADH and demonstrated that it is applicable for one-step synthesis of d-amino acids, such as d-leucine and d-isoleucine. As the next step, the creation of an enzyme exhibiting different substrate specificity and higher catalytic efficiency is a key to the further development of d-amino acid production. In this study, we succeeded in creating a novel mutant exhibiting extremely high catalytic activity for phenylpyruvate amination. Structural insight into the mutant will be useful for further improvement of DAADHs.
Keywords: NADP; Ureibacillus thermosphaericus; d-amino acid; d-phenylalanine; dehydrogenases; meso-diaminopimelate; phenylpyruvate.
Copyright © 2017 American Society for Microbiology.
Figures
Similar articles
-
Artificial Thermostable D-Amino Acid Dehydrogenase: Creation and Application.Front Microbiol. 2018 Aug 3;9:1760. doi: 10.3389/fmicb.2018.01760. eCollection 2018. Front Microbiol. 2018. PMID: 30123202 Free PMC article. Review.
-
Creation of a thermostable NADP⁺-dependent D-amino acid dehydrogenase from Ureibacillus thermosphaericus strain A1 meso-diaminopimelate dehydrogenase by site-directed mutagenesis.Biotechnol Lett. 2012 Sep;34(9):1693-9. doi: 10.1007/s10529-012-0952-1. Epub 2012 May 22. Biotechnol Lett. 2012. PMID: 22618239
-
Structural insight into the thermostable NADP(+)-dependent meso-diaminopimelate dehydrogenase from Ureibacillus thermosphaericus.Acta Crystallogr D Biol Crystallogr. 2015 May;71(Pt 5):1136-46. doi: 10.1107/S1399004715003673. Epub 2015 Apr 24. Acta Crystallogr D Biol Crystallogr. 2015. PMID: 25945579
-
Essential role of amino acid position 71 in substrate preference by meso-diaminopimelate dehydrogenase from Symbiobacterium thermophilum IAM14863.Enzyme Microb Technol. 2018 Apr;111:57-62. doi: 10.1016/j.enzmictec.2018.01.001. Epub 2018 Jan 3. Enzyme Microb Technol. 2018. PMID: 29421037
-
Creation of thermostable l-tryptophan dehydrogenase by protein engineering and its application for l-tryptophan quantification.Anal Biochem. 2019 Aug 15;579:57-63. doi: 10.1016/j.ab.2019.05.010. Epub 2019 May 14. Anal Biochem. 2019. PMID: 31100220 Review.
Cited by
-
Rational Design of Meso-Diaminopimelate Dehydrogenase with Enhanced Reductive Amination Activity for Efficient Production of d-p-Hydroxyphenylglycine.Appl Environ Microbiol. 2023 May 31;89(5):e0010923. doi: 10.1128/aem.00109-23. Epub 2023 Apr 18. Appl Environ Microbiol. 2023. PMID: 37070978 Free PMC article.
-
Diaminopimelic Acid Metabolism by Pseudomonadota in the Ocean.Microbiol Spectr. 2022 Oct 26;10(5):e0069122. doi: 10.1128/spectrum.00691-22. Epub 2022 Aug 30. Microbiol Spectr. 2022. PMID: 36040174 Free PMC article.
-
Amino Acid Chirality: Stereospecific Conversion and Physiological Implications.ACS Omega. 2024 Jan 26;9(5):5084-5099. doi: 10.1021/acsomega.3c08305. eCollection 2024 Feb 6. ACS Omega. 2024. PMID: 38343938 Free PMC article. Review.
-
Advances in Enzymatic Synthesis of D-Amino Acids.Int J Mol Sci. 2020 May 1;21(9):3206. doi: 10.3390/ijms21093206. Int J Mol Sci. 2020. PMID: 32369969 Free PMC article. Review.
-
Artificial Thermostable D-Amino Acid Dehydrogenase: Creation and Application.Front Microbiol. 2018 Aug 3;9:1760. doi: 10.3389/fmicb.2018.01760. eCollection 2018. Front Microbiol. 2018. PMID: 30123202 Free PMC article. Review.
References
-
- Akita H, Doi K, Kawarabayasi Y, Ohshima T. 2012. Creation of a thermostable NADP+-dependent d-amino acid dehydrogenase from Ureibacillus thermosphaericus strain A1 meso-diaminopimelate dehydrogenase by site-directed mutagenesis. Biotechnol Lett 34:1693–1699. doi:10.1007/s10529-012-0952-1. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
