

Mechanisms for Pseudoalteromonas piscicida-Induced Killing of Vibrios and Other Bacterial Pathogens
- PMID: 28363962
- PMCID: PMC5440704
- DOI: 10.1128/AEM.00175-17
Mechanisms for Pseudoalteromonas piscicida-Induced Killing of Vibrios and Other Bacterial Pathogens
Abstract
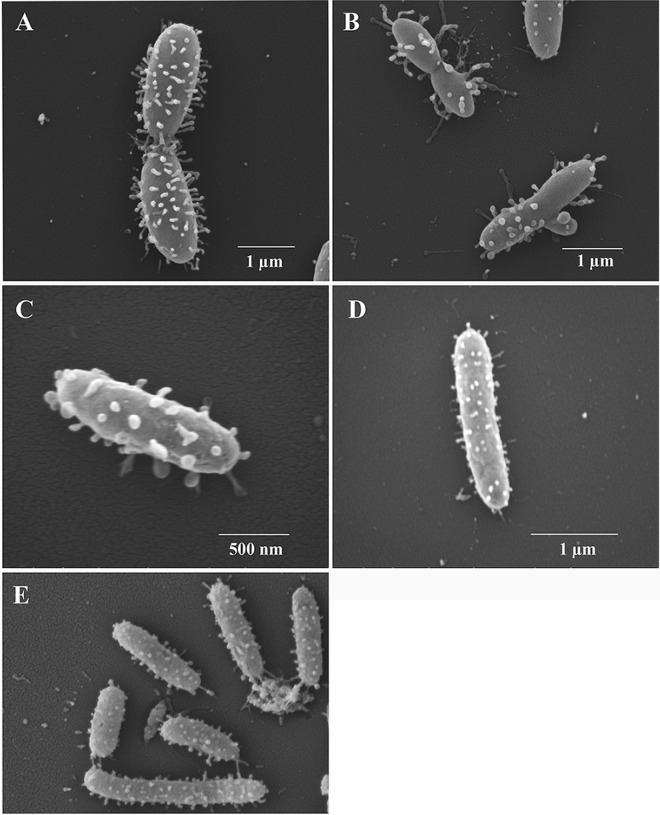
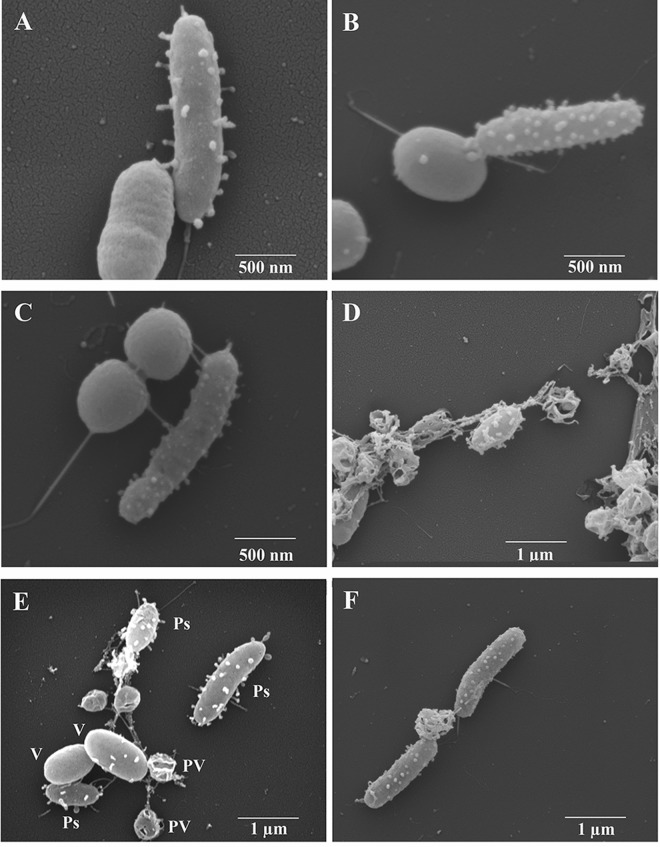
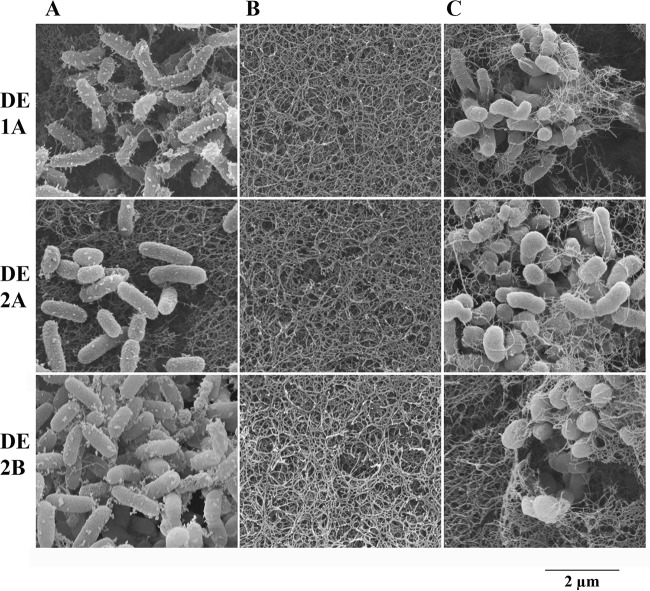
Pseudoalteromonas piscicida is a Gram-negative gammaproteobacterium found in the marine environment. Three strains of pigmented P. piscicida were isolated from seawater and partially characterized by inhibition studies, electron microscopy, and analysis for proteolytic enzymes. Growth inhibition and death occurred around colonies of P. piscicida on lawns of the naturally occurring marine pathogens Vibrio vulnificus, Vibrio parahaemolyticus, Vibrio cholerae, Photobacterium damselae, and Shewanella algae Inhibition also occurred on lawns of Staphylococcus aureus but not on Escherichia coli O157:H7 or Salmonella enterica serovar Typhimurium. Inhibition was not pH associated, but it may have been related to the secretion of a cysteine protease with strong activity, as detected with a synthetic fluorogenic substrate. This diffusible enzyme was secreted from all three P. piscicida strains. Direct overlay of the Pseudoalteromonas colonies with synthetic fluorogenic substrates demonstrated the activity of two aminopeptidase Bs, a trypsin-like serine protease, and enzymes reactive against substrates for cathepsin G-like and caspase 1-like proteases. In seawater cultures, scanning electron microscopy revealed numerous vesicles tethered to the outer surface of P. piscicida and a novel mechanism of direct transfer of these vesicles to V. parahaemolyticus Vesicles digested holes in V. parahaemolyticus cells, while the P. piscicida congregated around the vibrios in a predatory fashion. This transfer of vesicles and vesicle-associated digestion of holes were not observed in other bacteria, suggesting that vesicle binding may be mediated by host-specific receptors. In conclusion, we show two mechanisms by which P. piscicida inhibits and/or kills competing bacteria, involving the secretion of antimicrobial substances and the direct transfer of digestive vesicles to competing bacteria.IMPORTANCEPseudoalteromonas species are widespread in nature and reduce competing microflora by the production of antimicrobial compounds. We isolated three strains of P. piscicida and characterized secreted and cell-associated proteolytic enzymes, which may have antimicrobial properties. We identified a second method by which P. piscicida kills V. parahaemolyticus It involves the direct transfer of apparently lytic vesicles from the surface of the Pseudoalteromonas strains to the surface of Vibrio cells, with subsequent digestion of holes in the Vibrio cell walls. Enzymes associated with these vesicles are likely responsible for the digestion of holes in the cell walls. Pseudoalteromonas piscicida has potential applications in aquaculture and food safety, in control of the formation of biofilms in the environment, and in food processing. These findings may facilitate the probiotic use of P. piscicida to inactivate pathogens and may lead to the isolation of enzymes and other antimicrobial compounds of pharmacological value.
Keywords: Pseudoalteromonas piscicida; Vibrio cholerae; Vibrio parahaemolyticus; Vibrio vulnificus; bacterial inhibition; enzymes; lytic vesicles; predatory bacteria; probiotics; proteases.
Copyright © 2017 American Society for Microbiology.
Figures



References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
