

Impairment of human neural crest cell migration by prolonged exposure to interferon-beta
- PMID: 28365849
- PMCID: PMC5608792
- DOI: 10.1007/s00204-017-1966-1
Impairment of human neural crest cell migration by prolonged exposure to interferon-beta
Erratum in
-
Letter to the editor.Arch Toxicol. 2019 Jun;93(6):1771. doi: 10.1007/s00204-019-02472-z. Epub 2019 May 13. Arch Toxicol. 2019. PMID: 31087121 Free PMC article. No abstract available.
Abstract
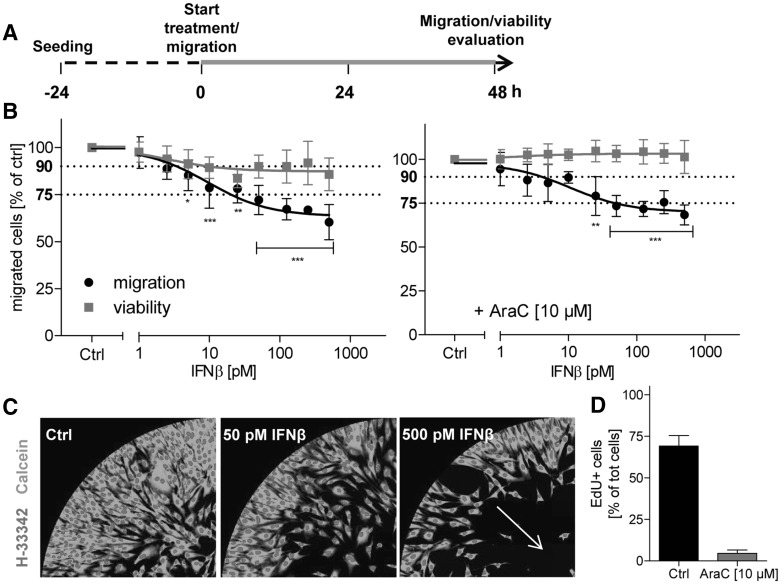
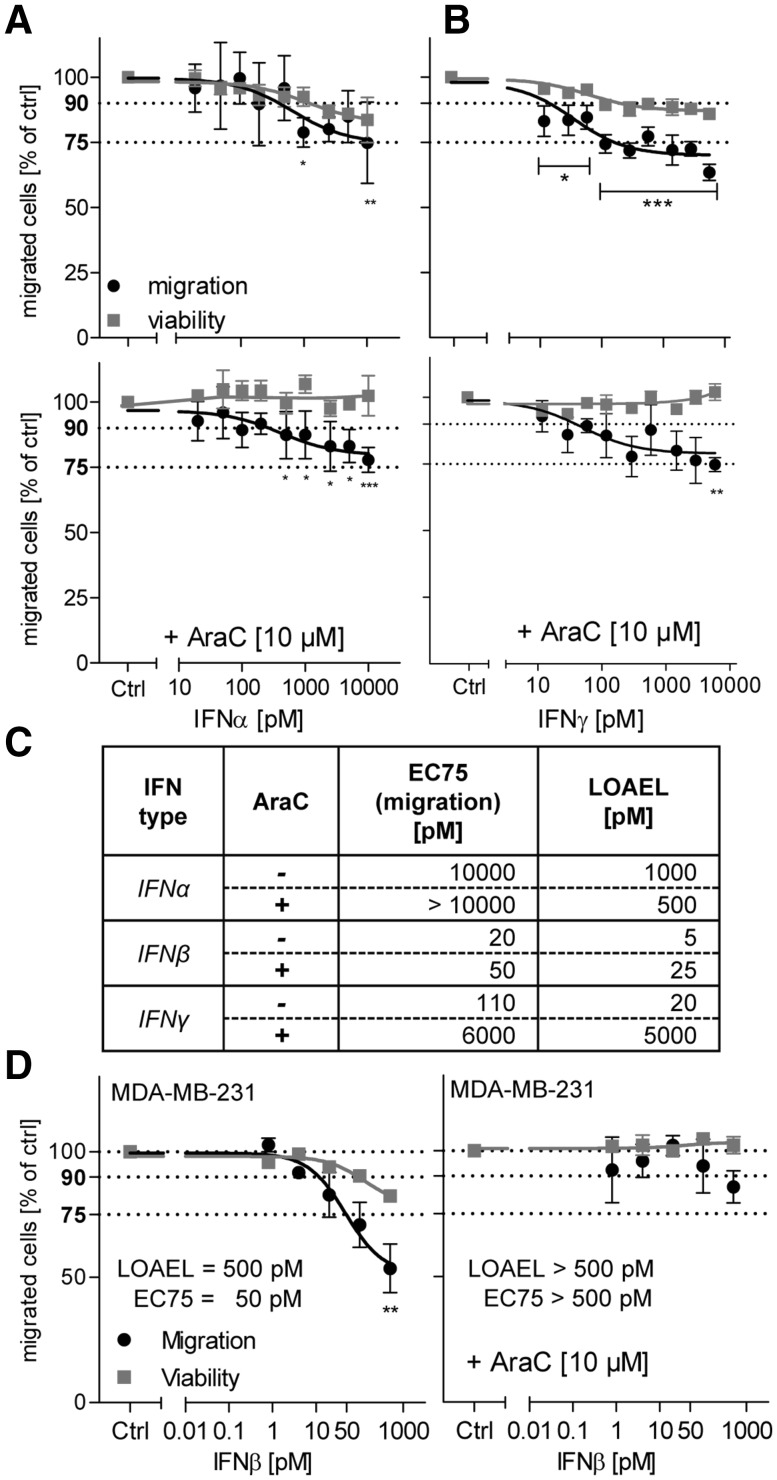
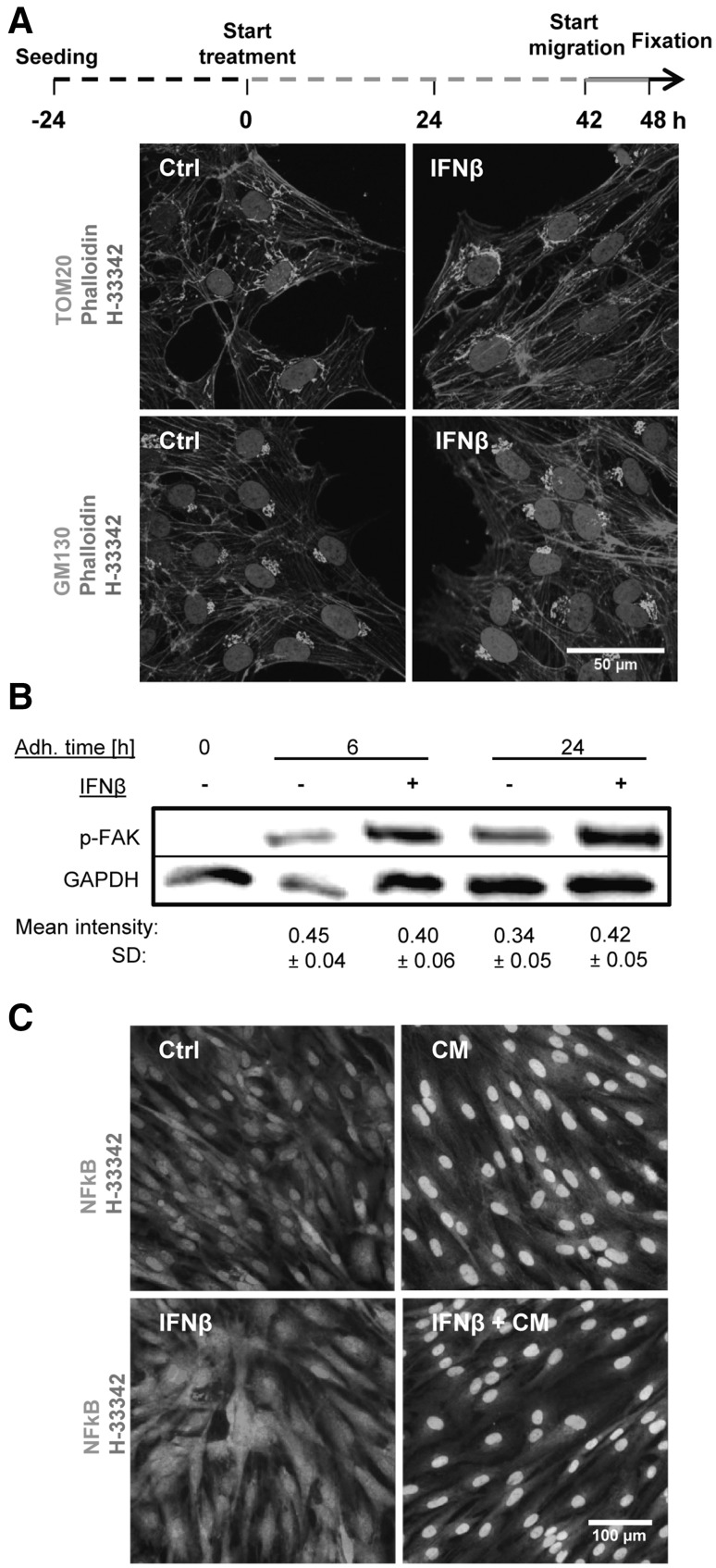
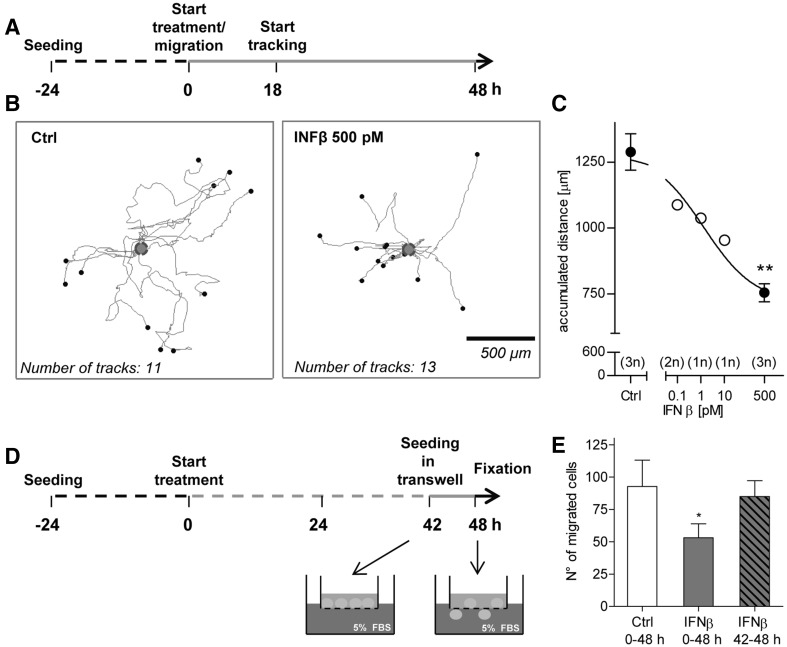
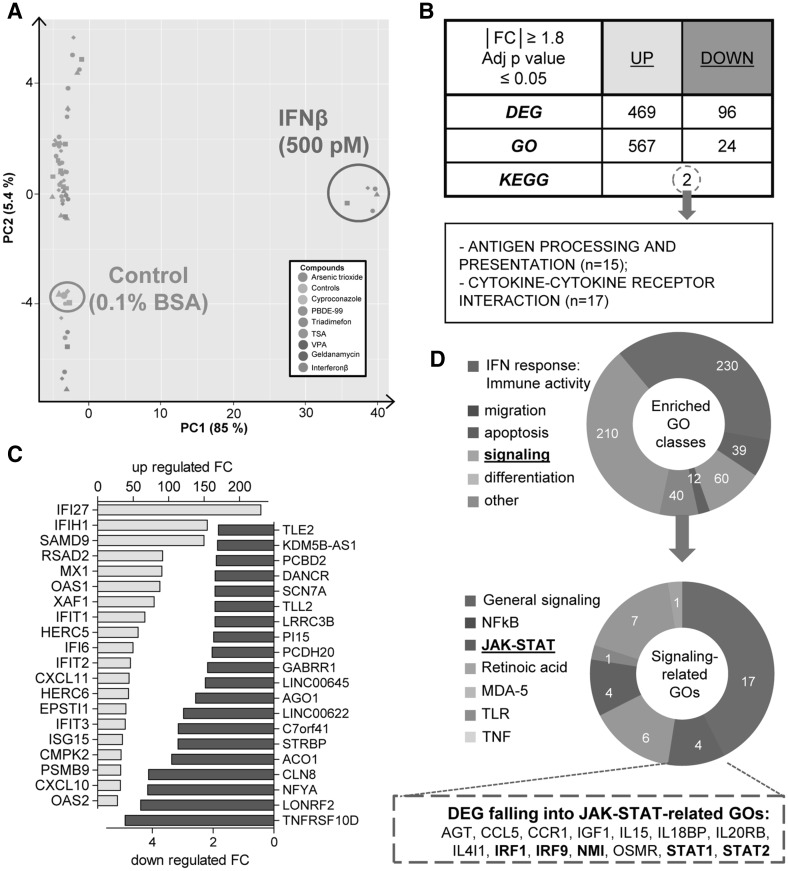
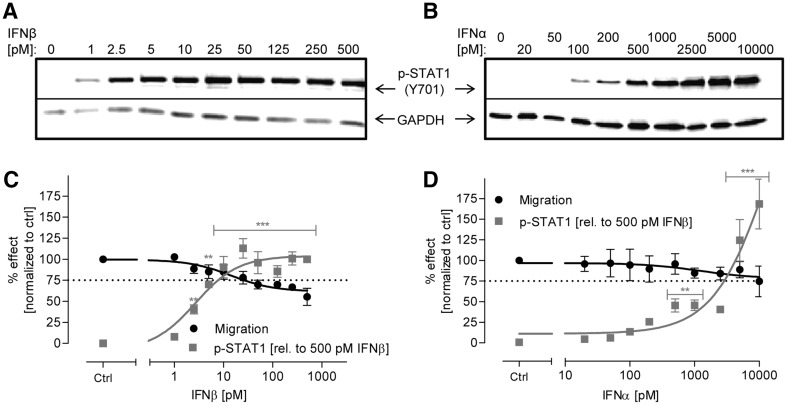
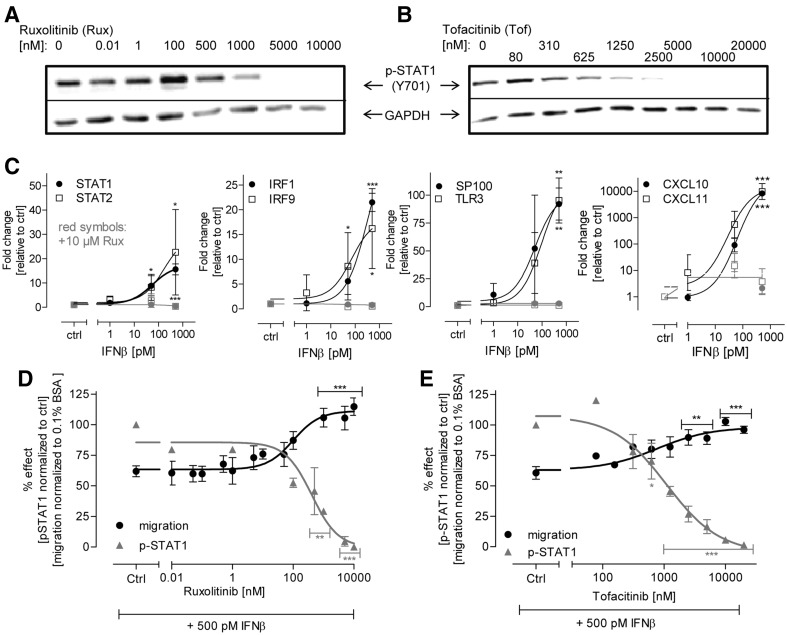
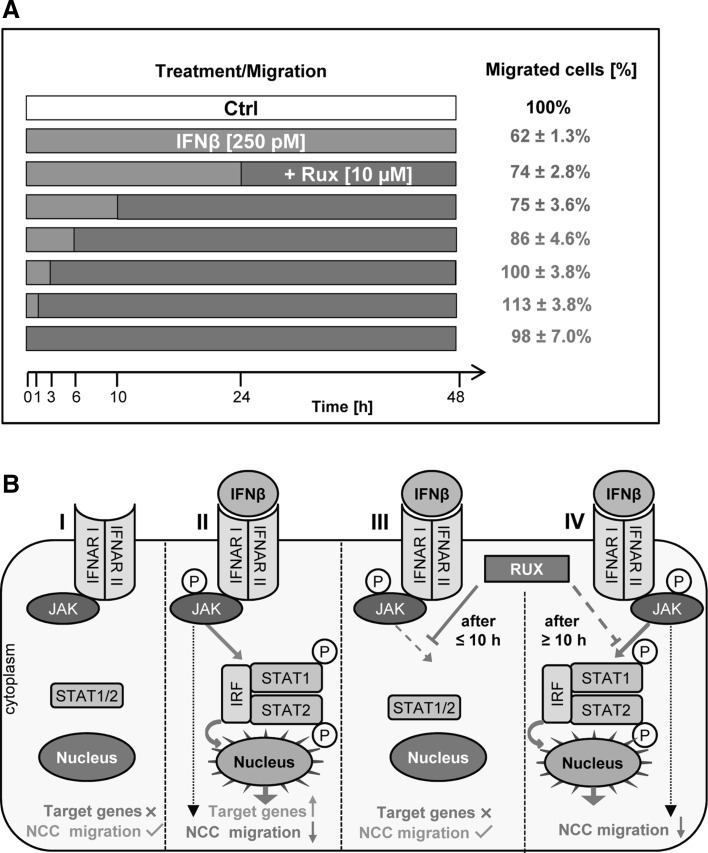
Human cell-based toxicological assays have been used successfully to detect known toxicants, and to distinguish them from negative controls. However, there is at present little experience on how to deal with hits from screens of compounds with yet unknown hazard. As a case study to this issue, we characterized human interferon-beta (IFNβ) as potential developmental toxicant affecting neural crest cells (NCC). The protein was identified as a hit during a screen of clinically used drugs in the 'migration inhibition of neural crest' (MINC) assay. Concentration-response studies in the MINC combined with immunocytochemistry and mRNA quantification of cellular markers showed that IFNβ inhibited NCC migration at concentrations as low as 20 pM. The effective concentrations found here correspond to levels found in human plasma, and they were neither cytostatic nor cytotoxic nor did they did they affect the differentiation state and overall phenotype of NCC. Data from two other migration assays confirmed that picomolar concentration of IFNβ reduced the motility of NCC, while other interferons were less potent. The activation of JAK kinase by IFNβ, as suggested by bioinformatics analysis of the transcriptome changes, was confirmed by biochemical methods. The degree and duration of pathway activation correlated with the extent of migration inhibition, and pharmacological block of this signaling pathway before, or up to 6 h after exposure to the cytokine prevented the effects of IFNβ on migration. Thus, the reduction of vital functions of human NCC is a hitherto unknown potential hazard of endogenous or pharmacologically applied interferons.
Keywords: Cell migration; Developmental toxicity; Interferons; JAK-STAT pathway; Neural crest.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Alam JMA, Scaramucci J, Jones W, Rogge M. Pharmacokinetics and pharmacodynamics of interferon Beta-la (IFNβ-1a) in healthy volunteers after intravenous, subcutaneous or intramuscular administration. Clin Drug Investig. 1997;14:35. doi: 10.2165/00044011-199714010-00005. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
