

Transcriptomics Indicates Active and Passive Metronidazole Resistance Mechanisms in Three Seminal Giardia Lines
- PMID: 28367140
- PMCID: PMC5355454
- DOI: 10.3389/fmicb.2017.00398
Transcriptomics Indicates Active and Passive Metronidazole Resistance Mechanisms in Three Seminal Giardia Lines
Abstract
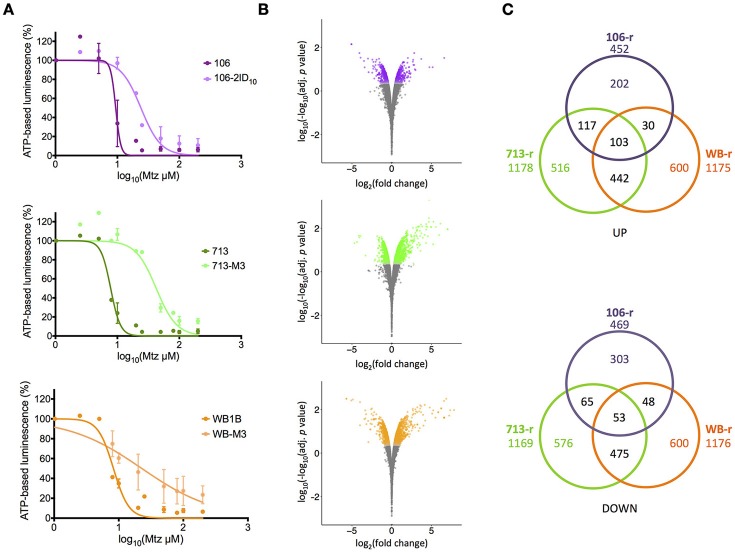
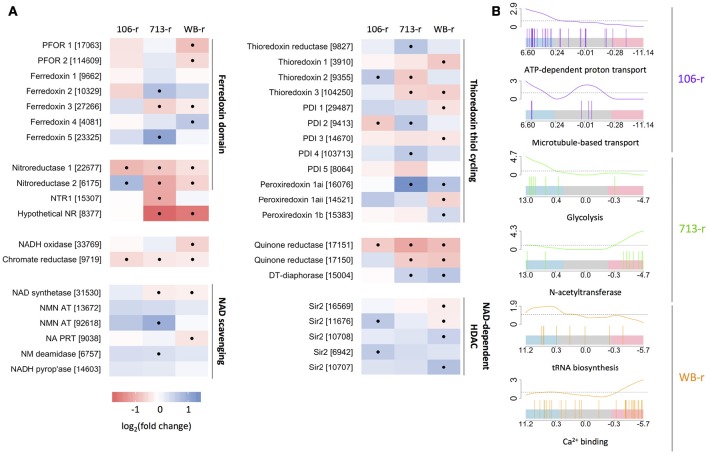
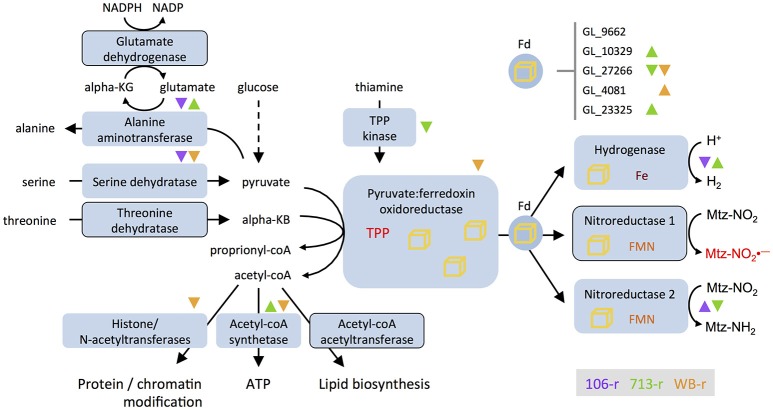
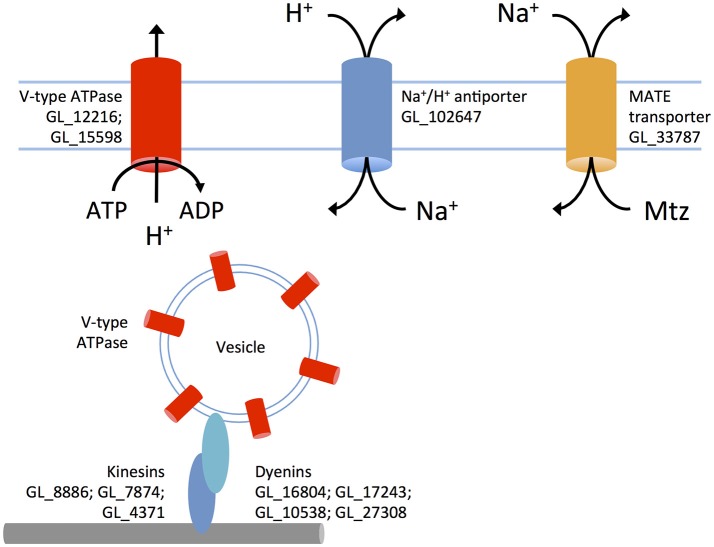
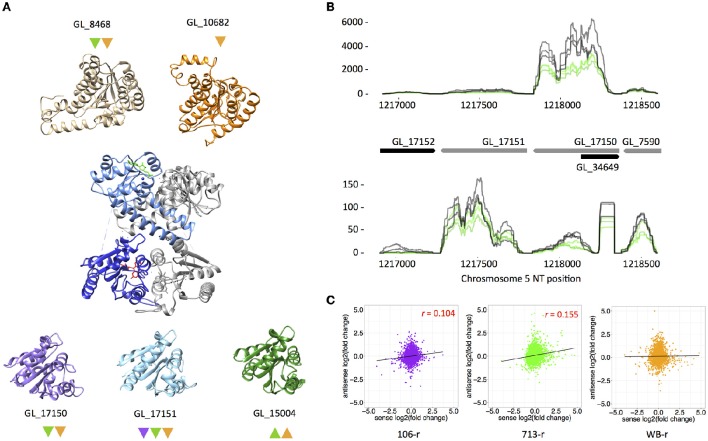
Giardia duodenalis is an intestinal parasite that causes 200-300 million episodes of diarrhoea annually. Metronidazole (Mtz) is a front-line anti-giardial, but treatment failure is common and clinical resistance has been demonstrated. Mtz is thought to be activated within the parasite by oxidoreductase enzymes, and to kill by causing oxidative damage. In G. duodenalis, Mtz resistance involves active and passive mechanisms. Relatively low activity of iron-sulfur binding proteins, namely pyruvate:ferredoxin oxidoreductase (PFOR), ferredoxins, and nitroreductase-1, enable resistant cells to passively avoid Mtz activation. Additionally, low expression of oxygen-detoxification enzymes can allow passive (non-enzymatic) Mtz detoxification via futile redox cycling. In contrast, active resistance mechanisms include complete enzymatic detoxification of the pro-drug by nitroreductase-2 and enhanced repair of oxidized biomolecules via thioredoxin-dependent antioxidant enzymes. Molecular resistance mechanisms may be largely founded on reversible transcriptional changes, as some resistant lines revert to drug sensitivity during drug-free culture in vitro, or passage through the life cycle. To comprehensively characterize these changes, we undertook strand-specific RNA sequencing of three laboratory-derived Mtz-resistant lines, 106-2ID10, 713-M3, and WB-M3, and compared transcription relative to their susceptible parents. Common up-regulated genes encoded variant-specific surface proteins (VSPs), a high cysteine membrane protein, calcium and zinc channels, a Mad-2 cell cycle regulator and a putative fatty acid α-oxidase. Down-regulated genes included nitroreductase-1, putative chromate and quinone reductases, and numerous genes that act proximal to PFOR. Transcriptional changes in 106-2ID10 diverged from those in 713-r and WB-r (r ≤ 0.2), which were more similar to each other (r = 0.47). In 106-2ID10, a nonsense mutation in nitroreductase-1 transcripts could enhance passive resistance whereas increased transcription of nitroreductase-2, and a MATE transmembrane pump system, suggest active drug detoxification and efflux, respectively. By contrast, transcriptional changes in 713-M3 and WB-M3 indicated a higher oxidative stress load, attributed to Mtz- and oxygen-derived radicals, respectively. Quantitative comparisons of orthologous gene transcription between Mtz-resistant G. duodenalis and Trichomonas vaginalis, a closely related parasite, revealed changes in transcripts encoding peroxidases, heat shock proteins, and FMN-binding oxidoreductases, as prominent correlates of resistance. This work provides deep insight into Mtz-resistant G. duodenalis, and illuminates resistance-associated features across parasitic species.
Keywords: Giardia; RNA sequencing (RNA-Seq); Trichomonas; messenger RNA; metronidazole resistance; oxidoreductases; single nucleotide polymorphism.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources