

Ionic currents influencing spontaneous firing and pacemaker frequency in dopamine neurons of the ventrolateral periaqueductal gray and dorsal raphe nucleus (vlPAG/DRN): A voltage-clamp and computational modelling study
- PMID: 28367595
- PMCID: PMC5403876
- DOI: 10.1007/s10827-017-0641-0
Ionic currents influencing spontaneous firing and pacemaker frequency in dopamine neurons of the ventrolateral periaqueductal gray and dorsal raphe nucleus (vlPAG/DRN): A voltage-clamp and computational modelling study
Abstract
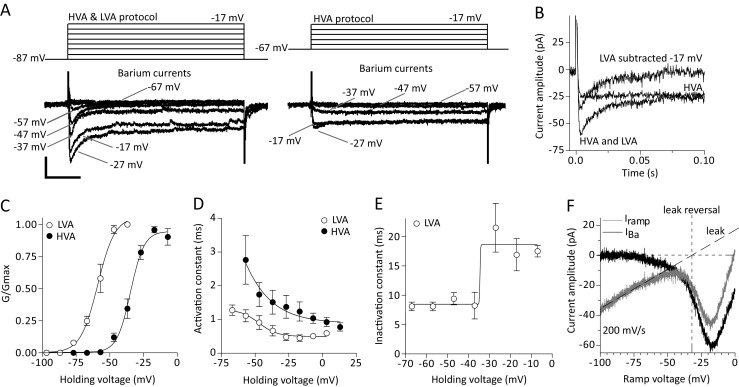
Dopamine (DA) neurons of the ventrolateral periaqueductal gray (vlPAG) and dorsal raphe nucleus (DRN) fire spontaneous action potentials (APs) at slow, regular patterns in vitro but a detailed account of their intrinsic membrane properties responsible for spontaneous firing is currently lacking. To resolve this, we performed a voltage-clamp electrophysiological study in brain slices to describe their major ionic currents and then constructed a computer model and used simulations to understand the mechanisms behind autorhythmicity in silico. We found that vlPAG/DRN DA neurons exhibit a number of voltage-dependent currents activating in the subthreshold range including, a hyperpolarization-activated cation current (IH), a transient, A-type, potassium current (IA), a background, 'persistent' (INaP) sodium current and a transient, low voltage activated (LVA) calcium current (ICaLVA). Brain slice pharmacology, in good agreement with computer simulations, showed that spontaneous firing occurred independently of IH, IA or calcium currents. In contrast, when blocking sodium currents, spontaneous firing ceased and a stable, non-oscillating membrane potential below AP threshold was attained. Using the DA neuron model we further show that calcium currents exhibit little activation (compared to sodium) during the interspike interval (ISI) repolarization while, any individual potassium current alone, whose blockade positively modulated AP firing frequency, is not required for spontaneous firing. Instead, blockade of a number of potassium currents simultaneously is necessary to eliminate autorhythmicity. Repolarization during ISI is mediated initially via the deactivation of the delayed rectifier potassium current, while a sodium background 'persistent' current is essentially indispensable for autorhythmicity by driving repolarization towards AP threshold.
Keywords: Autorhythmicity; Delayed rectifier; Depolarization block; Electrophysiology; Persistent sodium current.
Conflict of interest statement
Conflict of interest
The authors declare that they have no conflict of interest.
Software availability
The model and simulation experiments described herein have been conducted using free access software (NEURON,
Figures









References
-
- Brevi S, de Curtis M, Magistretti J. Pharmacological and biophysical characterization of voltage-gated calcium currents in the endopiriform nucleus of the guinea pig. Journal of Neurophysiology. 2001;85:2076–2087. - PubMed
-
- Brown, D. A., & Adams, P. R. (1980) Muscarinic suppression of a novel voltage-sensitive K+ current in a vertebrate neurone. Nature, 283, 673–676. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
