

Macrocycle peptides delineate locked-open inhibition mechanism for microorganism phosphoglycerate mutases
- PMID: 28368002
- PMCID: PMC5382265
- DOI: 10.1038/ncomms14932
Macrocycle peptides delineate locked-open inhibition mechanism for microorganism phosphoglycerate mutases
Abstract
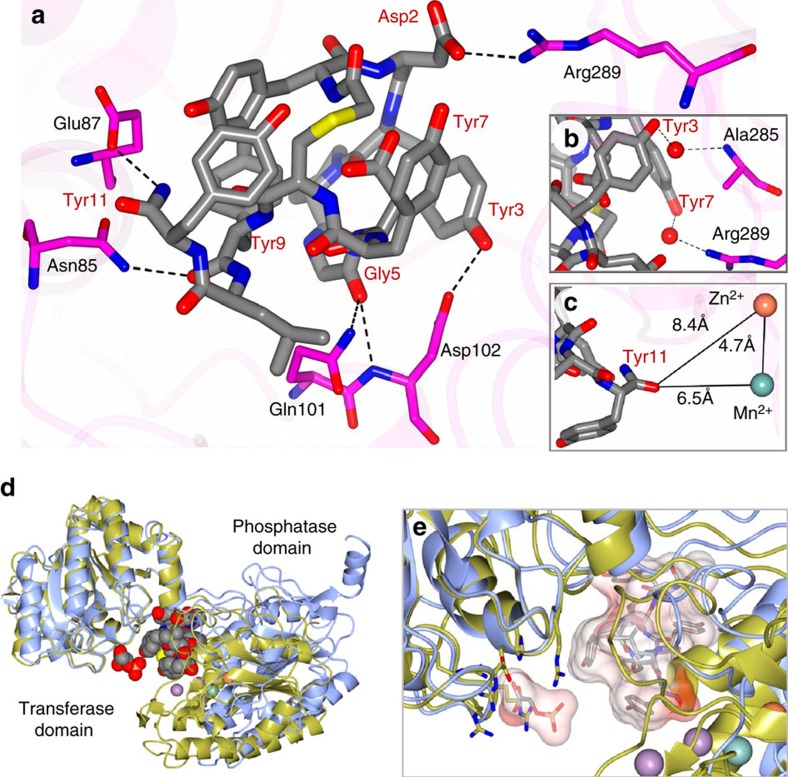
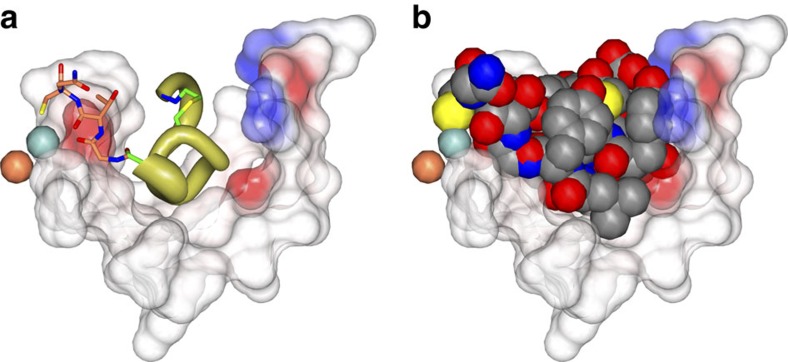
Glycolytic interconversion of phosphoglycerate isomers is catalysed in numerous pathogenic microorganisms by a cofactor-independent mutase (iPGM) structurally distinct from the mammalian cofactor-dependent (dPGM) isozyme. The iPGM active site dynamically assembles through substrate-triggered movement of phosphatase and transferase domains creating a solvent inaccessible cavity. Here we identify alternate ligand binding regions using nematode iPGM to select and enrich lariat-like ligands from an mRNA-display macrocyclic peptide library containing >1012 members. Functional analysis of the ligands, named ipglycermides, demonstrates sub-nanomolar inhibition of iPGM with complete selectivity over dPGM. The crystal structure of an iPGM macrocyclic peptide complex illuminated an allosteric, locked-open inhibition mechanism placing the cyclic peptide at the bi-domain interface. This binding mode aligns the pendant lariat cysteine thiolate for coordination with the iPGM transition metal ion cluster. The extended charged, hydrophilic binding surface interaction rationalizes the persistent challenges these enzymes have presented to small-molecule screening efforts highlighting the important roles of macrocyclic peptides in expanding chemical diversity for ligand discovery.
Conflict of interest statement
The authors declare no competing financial interests.
Figures




References
-
- World Health Organization. The World Health Report: 2004: Changing History (2004).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
