

Structure of acid deoxyribonuclease
- PMID: 28369538
- PMCID: PMC5449587
- DOI: 10.1093/nar/gkx222
Structure of acid deoxyribonuclease
Abstract
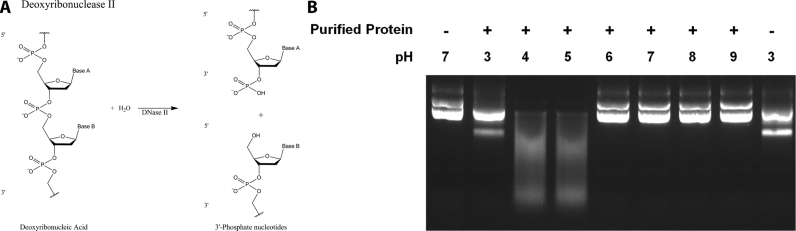
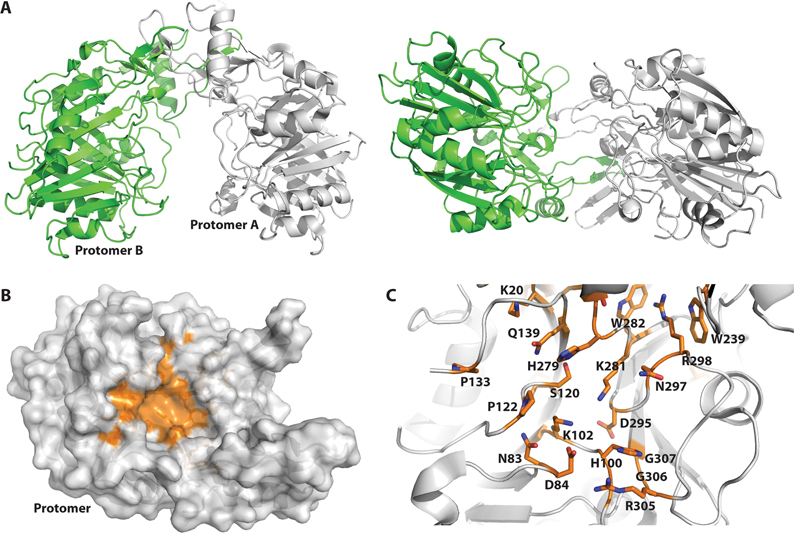
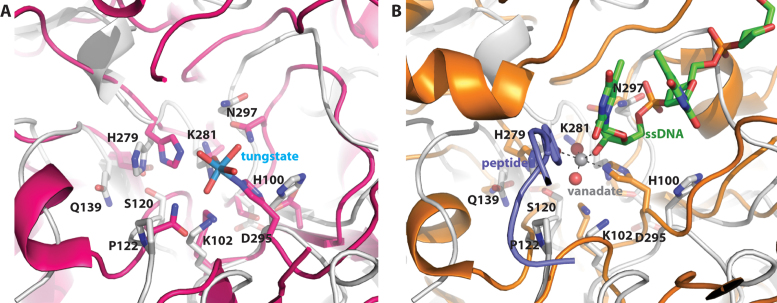
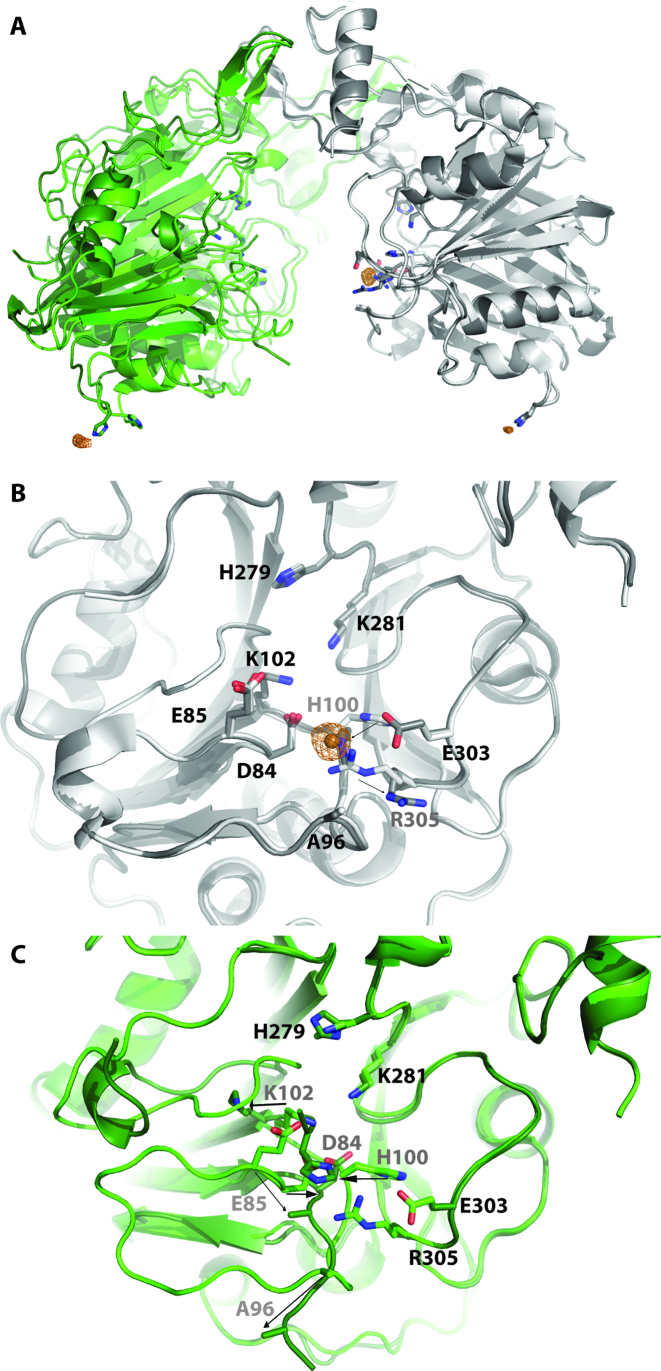
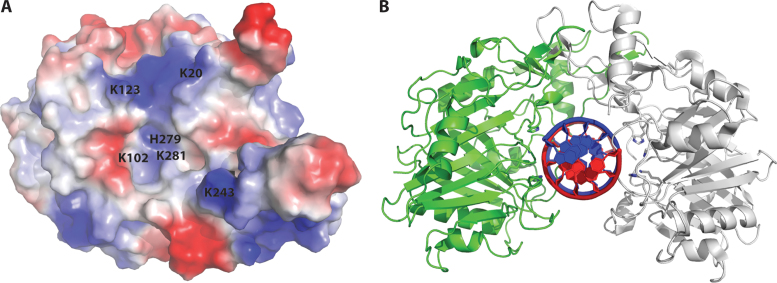
Deoxyribonuclease II (DNase II) is also known as acid deoxyribonuclease because it has optimal activity at the low pH environment of lysosomes where it is typically found in higher eukaryotes. Interestingly, DNase II has also been identified in a few genera of bacteria and is believed to have arisen via horizontal transfer. Here, we demonstrate that recombinant Burkholderia thailandensis DNase II is highly active at low pH in the absence of divalent metal ions, similar to eukaryotic DNase II. The crystal structure of B. thailandensis DNase II shows a dimeric quaternary structure which appears capable of binding double-stranded DNA. Each monomer of B. thailandensis DNase II exhibits a similar overall fold as phospholipase D (PLD), phosphatidylserine synthase (PSS) and tyrosyl-DNA phosphodiesterase (TDP), and conserved catalytic residues imply a similar mechanism. The structural and biochemical data presented here provide insights into the atomic structure and catalytic mechanism of DNase II.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Evans C.J., Aguilera R.J.. DNase II: genes, enzymes and function. Gene. 2003; 322:1–15. - PubMed
-
- Steitz T.A. A mechanism for all polymerases. Nature. 1998; 391:231–232. - PubMed
-
- Yamanaka M., Tsubota Y., Anai M., Ishimatsu K., Okumura M., Katsuki S., Takagi Y.. Purification and properties of acid deoxyribonucleases of human gastric mucosa and cervix uteri. J. Biol. Chem. 1974; 249:3884–3889. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
